What does npm exec do? What is the difference between «npm exec» and «npx»?
What are the building blocks of OWL ontologies?
Learn more about «RDF star», «SPARQL star», «Turtle star», «JSON-LD star», «Linked Data star», and «Semantic Web star».
The Hadamard gate is one of the simplest quantum gates which acts on a single qubit.
Learn more about the bra–ket notation.
Progressive Cactus is an evolution of the Cactus multiple genome alignment protocol that uses the progressive alignment strategy.
The Human Genome Project is an ambitious project which is still underway.
What are SVMs (support vector machines)?
Find out more in Eckher’s article about TensorFlow.js and linear regression.
On the importance of centralised metadata registries at companies like Uber.
Facebook’s Nemo is a new custom-built platform for internal data discovery. Learn more about Facebook’s Nemo.
What is Data Commons (datacommons.org)? Read Eckher’s introduction to Data Commons (datacommons.org) to learn more about the open knowledge graph built from thousands of public datasets.
Learn more about how Bayer uses semantic web technologies for corporate asset management and why it enables the FAIR data in the corporate environment.
An introduction to WikiPathways by Eckher is an overview of the collaboratively edited structured biological pathway database that discusses the history of the project, applications of the open dataset, and ways to access the data programmatically.
Eckher’s article about question answering explains how question answering helps extract information from unstructured data and why it will become a go-to NLP technology for the enterprise.
Read more about how document understanding AI works, what its industry use cases are, and which cloud providers offer this technology as a service.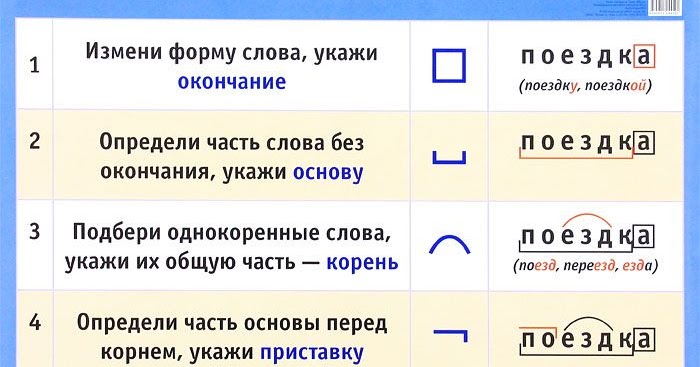
Lexemes are Wikidata’s new type of entity used for storing lexicographical information. The article explains the structure of Wikidata lexemes and ways to access the data, and discusses the applications of the linked lexicographical dataset.
The guide to exploring linked COVID-19 datasets describes the existing RDF data sources and ways to query them using SPARQL. Such linked data sources are easy to interrogate and augment with external data, enabling more comprehensive analysis of the pandemic both in New Zealand and internationally.
The introduction to the Gene Ontology graph published by Eckher outlines the structure of the GO RDF model and shows how the GO graph can be queried using SPARQL.
The overview of the Nobel Prize dataset published by Eckher demonstrates the power of Linked Data and demonstrates how linked datasets can be queried using SPARQL. Use SPARQL federation to combine the Nobel Prize dataset with DBPedia.
Learn why federated queries are an incredibly useful feature of SPARQL.
What are the best online Arabic dictionaries?
How to pronounce numbers in Arabic?
List of months in Maori.
Days of the week in Maori.
The list of country names in Tongan.
The list of IPA symbols.
What are the named entities?
What is computational linguistics?
Learn how to use the built-in React hooks.
Learn how to use language codes in HTML.
Learn about SSML.
Browse the list of useful UX resources from Google.
Where to find the emoji SVG sources?.
What is Wikidata?
What’s the correct markup for multilingual websites?
How to use custom JSX/HTML attributes in TypeScript?
Learn more about event-driven architecture.
Where to find the list of all emojis?
How to embed YouTube into Markdown?
What is the Google Knowledge Graph?
Learn SPARQL.
Explore the list of coronavirus (COVID-19) resources for bioinformaticians and data science researchers.
Sequence logos visualize protein and nucleic acid motifs and patterns identified through multiple sequence alignment. They are commonly used widely to represent transcription factor binding sites and other conserved DNA and RNA sequences. Protein sequence logos are also useful for illustrating various biological properties of proteins. Create a sequence logo with Sequence Logo. Paste your multiple sequence alignment and the sequence logo is generated automatically. Use the sequence logo maker to easily create vector sequence logo graphs. Please refer to the Sequence Logo manual for the sequence logo parameters and configuration. Sequence Logo supports multiple color schemes and download formats.
Sequence Logo is a web-based sequence logo generator. Sequence Logo generates sequence logo diagrams for proteins and nucleic acids. Sequence logos represent patterns found within multiple sequence alignments. They consist of stacks of letters, each representing a position in the sequence alignment. Sequence Logo analyzes the sequence data inside the user’s web browser and does not store or transmit the alignment data via servers.
Te Reo Maps is an online interactive Maori mapping service. All labels in Te Reo Maps are in Maori, making it the first interactive Maori map. Te Reo Maps is the world map, with all countries and territories translated into Maori. Please refer to the list of countries in Maori for the Maori translations of country names. The list includes all UN members and sovereign territories.
Phonetically is a web-based text-to-IPA transformer. Phonetically uses machine learning to predict the pronunciation of English words and transcribes them using IPA.
Punycode.org is a tool for converting Unicode-based internationalized domain names to ASCII-based Punycode encodings. Use punycode.org to quickly convert Unicode to Punycode and vice versa. Internationalized domains names are a new web standard that allows using non-ASCII characters in web domain names.
Use punycode.org to quickly convert Unicode to Punycode and vice versa. Internationalized domains names are a new web standard that allows using non-ASCII characters in web domain names.
My Sequences is an online platform for storing and analyzing personal sequence data. My Sequences allows you to upload your genome sequences and discover insights and patterns in your own DNA.
Словообразовательный словарь «Морфема» дает представление о морфемной структуре слов русского языка и слов современной лексики. Для словообразовательного анализа представлены наиболее употребительные слова современного русского языка, их производные и словоформы. Словарь предназначен школьникам, студентам и преподавателям. Статья разбора слова «сладкоежка» по составу показывает, что это слово имеет два корня, соединительную гласную, суффикс и окончание. На странице также приведены слова, содержащие те же морфемы. Словарь «Морфема» включает в себя не только те слова, состав которых анализируется в процессе изучения предмета, но и множество других слов современного русского языка. Словарь адресован всем, кто хочет лучше понять структуру русского языка.
Разбор слова «кормушка» по составу.
Разбор слова «светить» по составу.
Разбор слова «сбоку» по составу.
Разбор слова «шиповник» по составу.
Разбор слова «народ» по составу.
Разбор слова «впервые» по составу.
Разбор слова «свежесть» по составу.
Разбор слова «издалека» по составу.
Разбор слова «лесной» по составу.
«Темная башня», «Теснота», «Леди Макбет» и «Ветреная река»
Девушка в беде в одном из главных российских дебютов последнего времени
Кавказ, 1998 год. Молодая Ила из Нальчика (очень интересная Дарья Жовнер) больше всего любит работать в автомастерской у отца и выглядит соответственно: одета в свитер и комбинезон на три размера больше. Она живет в традиционной еврейской семье, где все решения принимаются старшими, и наперекор им заводит отношения с кабардинцем — обычным парнем с автозаправки. Младший брат Илы Давид собирается жениться на Лее из другой еврейской семьи: сообщество собирается дома и радостно отмечает помолвку. На следующее утро мать получает куртку сына и номер телефона, по которому можно пообщаться с похитителями: Давида и Лею украл для выкупа кто-то из местных. В полицию обращаться нельзя — будет только хуже. На Илу начинают давить, чтобы она пожертвовала своей личной жизнью ради брата. В семейных спорах все чаще звучит аргумент: нужно выбирать кого-то «из своего племени». Какого племени Ила, она сама не знает.
Она живет в традиционной еврейской семье, где все решения принимаются старшими, и наперекор им заводит отношения с кабардинцем — обычным парнем с автозаправки. Младший брат Илы Давид собирается жениться на Лее из другой еврейской семьи: сообщество собирается дома и радостно отмечает помолвку. На следующее утро мать получает куртку сына и номер телефона, по которому можно пообщаться с похитителями: Давида и Лею украл для выкупа кто-то из местных. В полицию обращаться нельзя — будет только хуже. На Илу начинают давить, чтобы она пожертвовала своей личной жизнью ради брата. В семейных спорах все чаще звучит аргумент: нужно выбирать кого-то «из своего племени». Какого племени Ила, она сама не знает.
Фильм снимал Кантемир Балагов, 26-летний ученик Александра Сокурова — несколько лет назад один из главных режиссеров России открыл экспериментальный курс в Нальчике и в результате выпустил дюжину молодых авторов. Так появившийся практически из ниоткуда Балагов получил приз ФИПРЕССИ в Каннах, награду за лучший дебют в «Кинотавре» и почти единодушно покорил критиков. Параллельно с этим в родной режиссеру Кабардино-Балкарии фильм обещали не показывать.
«Теснота» — это не только кино, снятое очень молодым человеком, но и фильм не из привычного столичного контекста. Балагов работает с совершенно не звучащим в современном отечественном кино кавказским регионом, темой традиции и иерархии без кретинических стереотипов: вспомните последнего героя с Кавказа, который не был бы террористом, уборщиком, мафиози или торговцем. Сюжет «Тесноты» собран из реальных случаев похищений, часто происходивших в Кабардино-Балкарии и вообще на Кавказе: в 90-е еврейские семьи были довольно состоятельными, но держались особняком, и кража члена другого клана была распространенным способом заработать. Герои «Тесноты» выживают, как и большинство в 90-е, в постоянной обороне за свою идентичность. Песни Тани Булановой и группы «Вирус», спортивные костюмы, война в Чечне, горы фоном, клеенка на столе, разбитые дороги, халтура ради выживания — режиссер Кантемир Балагов хорошо понимает, помнит и любит места, о которых снимает.
В «Тесноте» вообще не существует государства, есть только сообщество и призрак войны, а маркировка «свой/чужой» обеспечивает простые правила жизни на каждый день. Именно поэтому одинокая женская фигура — стопроцентная маргиналка, безопасность и будущее которой никто не может гарантировать: Дарья Жовнер идеально играет человека, не ценного самого по себе. В «Тесноте» много грубых швов и упрощений, объяснимых для дебютной работы — слабые и сильные актеры играют слишком по-разному, цвет пытается объяснить (иногда даже разжевать) сюжет, а теснота из названия очень буквально реализована в каждой сцене. Тем не менее этот фильм по настроению и интимности — обещание большого будущего для исполнительницы главной роли и режиссера, чутких и внимательных к неоднозначности человека. В первом за долгое время российском дебюте о травмирующей семье проговариваются такие узнаваемые и необходимые для озвучивания послания, которые слышит каждый второй молодой человек на просторах нашей страны: «ты никто, пока не создашь семью», «какая разница, чего ты хочешь», «ищи человека своего круга», «ты ни на что не годишься». Без замаха на великое кино и огромный бюджет «Теснота» — здоровый и полезный пример рефлексии и самокритики над собой, близкими, поколением и друзьями, через который проживаются общие травмы, а распространенные явления называются своими именами. Отсутствие границ, давление, угрозы, манипуляции — в общем, теснота.
В ТЕСНОТЕ И… В ОБИДЕ
Вместе с комендантским патрулём главка корреспонденты газеты «Петровка, 38» побывали в ОМВД России по району Гагаринский Юго-Западного округа столицы.После инструктажа, который провели сотрудники Инспекции по личному составу, наш автомобиль направляется в один из самых престижных районов города. Знакомимся с сотрудниками патруля. Подполковник полиции Андрей Жолудев и его помощник, лейтенант полиции Алексей Бутенков, демонстрируют нам план-задание на проверку подразделения. В документе чётко прописано, с чем патрулю необходимо ознакомиться.
Начинается инспектирование с КПП. Сотрудник, охраняющий вход в райотдел, чётко представляется, записывает в журнал наши персональные данные и докладывает о визите проверяющих в дежурную часть. Через минуту на крыльце появляется начальник отдела подполковник полиции Андрей Яковлев. В дальнейшем он становится нашим «гидом» во время обхода территории отдела. Двор подразделения совсем небольшой. Размером он сопоставим с волейбольной площадкой. Оно и понятно: рядом Ленинский проспект. Тут плотная застройка, поэтому и земля здесь — на вес золота. Но весь двор заставлен опечатанными автомобилями. Мы насчитываем их двадцать три. Это не считая служебного автотранспорта. Теснота — жуткая. Невольно проносится мысль: а вдруг пожар? Что делать, ведь сгорят все машины. Не сумеют их вовремя эвакуировать. Интересуемся у подполковника полиции Яковлева, почему так много транспортных средств на столь микроскопическом клочке земли. Узнаём, что, все иномарки — задержаны и являются вещдоками. Многие из них задержаны подразделениями ГАИ или вневедомственной охраны. И доставлены сюда по территориальности. Более того, отдельные «железные кони» занимают драгоценную площадь годами, и не ясно, что с ними делать. Например, полуразбитая «Мазда», прячущаяся под тентом, была доставлена сюда более двух лет назад, после случившегося ДТП. Но хозяин её не торопится забирать. Неужели на территории округа не найдётся места для специализированной охраняемой стоянки, где можно было бы размещать подобные транспортные средства, а не загромождать и без того маленькую территорию райотдела? Впрочем, этот вопрос мы адресуем префектуре Юго-Западного административного округа.
Следующий этап нашей «экспедиции» — дежурная часть отдела. Помощник оперативного дежурного оказывается нештатным. Эти обязанности временно возложены на оперуполномоченного уголовного розыска лейтенанта полиции Дмитрия Пянзина. Как выяснилось, произошёл форс-мажор. Штатный помощник заболел. И начальник райотдела официальным приказом временно назначил сыщика на эту должность. Однако патрульные фиксируют нарушение: отсутствие у этого офицера бейджика. Забегая вперёд, сообщаем, что данное замечание было устранено в течение минут пятнадцати.
Однако патрульные фиксируют нарушение: отсутствие у этого офицера бейджика. Забегая вперёд, сообщаем, что данное замечание было устранено в течение минут пятнадцати.
Не обнаружили проверяющие в холле и информационных стендов для посетителей. Но и это объяснимо. На момент проверки в здании производился ремонт. Кстати, само сооружение отнюдь не «молодо». Построено оно почти сорок лет назад. Мы начинаем обход помещений. Возле двери одного из кабинетов отсутствует табличка, на которой должны быть указаны должность и фамилия хозяина-сотрудника. Впрочем, этот недостаток устраняется незамедлительно. Патрульные проверяют наличие служебных удостоверений у сотрудников, их внешний вид. Нарушений выявлено не было. Зато во дворе проверяющие фиксируют ещё два недостатка: в курилке не имеется средств пожаротушения, а над мусорными баками нет навеса. Однако, в целом, данный райотдел производит на нас положительное впечатление. И, судя по решимости его руководителя, все выявленные замечания будут устранены в ближайшее время.
Михаил СМИРНОВ,
фото Николая ГОРБИКОВА
Объявлен шорт-лист первой кинопремии «Восток – Запад. Золотая арка»
© Конфедерация союзов кинематографистовСегодня, 27 марта, в пресс-центре МИА «Россия сегодня» в Москве прошла пресс-конференция, посвященная учреждению и объявлению шорт-листа первой международной кинематографической премии «Восток – Запад. Золотая арка», учрежденной Конфедерацией союзов кинематографистов.
В пресс-конференции приняли участие председатель правления премии, кинодраматург Рустам Ибрагимбеков, арт-директор премии, кинокритик Андрей Плахов и pr-директор премии, кинокритик Константин Шавловский.
Кинопремия «Восток — Запад. Золотая арка» присуждается за достижения и открытия в области киноискусства лучшим авторам фильмов, произведенных в странах Восточной Европы и Западной Азии. Она призвана стимулировать и поощрять эти открытия, привлекать внимание к ярким художественным процессам, которые происходят на территории Евразии и которые часто остаются в тени более коммерческого, преимущественно англоязычного кинематографа.
География премии обширна, список стран-участниц насчитывает 32 страны, и в будущем он может расшириться. На сегодняшний день в него входят Азербайджан, Албания, Армения, Беларусь, Босния и Герцеговина, Болгария, Венгрия, Греция, Грузия, Израиль, Казахстан, Кипр, Кыргызстан, Латвия, Литва, Македония, Молдова, Польша, Россия, Румыния, Сербия, Словакия, Словения, Таджикистан, Туркменистан, Турция, Узбекистан, Украина, Финляндия, Хорватия, Чехия, Эстония.
В жюри премии «Восток — Запад. Золотая арка» 2017/2018 вошли 24 киноведа и кинокритика — ведущие специалисты по кинематографу стран Азии и Восточной Европы. С полным составом жюри, а также с лонг-листом премии, в который отборочная комиссия под председательством Андрея Плахова включила 32 фильма, можно ознакомиться на сайте премии.
В шорт-лист вошли 13 фильмов. Лидерами по количеству номинаций стали российские картины «Нелюбовь» Андрея Звягинцева (5 номинаций) и «Теснота» Кантемира Балагова (5 номинаций). По 4 номинации у фильмов «Вестерн» Валески Гризебах (Болгария, Германия, Австрия) и «Фокстрот» Самуэля Маоза (Израиль).
Шорт-лист премии «Восток — Запад. Золотая арка»:
Лучший актер второго плана
- Сакари Куосманен, «По ту сторону надежды», реж. Аки Каурисмяки (Финляндия)
- Алексей Серебряков, «Как Витька Чеснок вез Леху Штыря в дом инвалидов», реж. Александр Хант (Россия)
- Шерван Хаджи, «По ту сторону надежды», реж. Аки Каурисмяки (Финляндия)
Лучшая актриса второго плана
- Джованна Гаврилович, «Реквием для Миссис Джей», реж. Боян Вулетич (Сербия-Болгария- Республика Македония-Россия-Франция)
- Ольга Драгунова, «Теснота», реж.
 Кантемир Балагов (Россия)
Кантемир Балагов (Россия) - Сара Адлер, «Фокстрот», реж. Самуэль Маоз (Израиль)
 Кантемир Балагов (Россия)
Кантемир Балагов (Россия)Лучший актер
- Александр Яценко, «Аритмия», реж. Борис Хлебников (Россия)
- Богдан Думитраке, «Поророка», реж. Константин Попеску (Румыния-Франция)
- Майнхард Нойман, «Вестерн», реж. Валеска Гризебах (Болгария-Германия-Австрия)
Лучшая актриса
- Александра Борбей, «О теле и душе», реж. Ильдико Эньеди (Венгрия)
- Дарья Жовнер, «Теснота», реж. Кантемир Балагов (Россия)
- Миряна Каранович, «Реквием для Миссис Джей», реж. Боян Вулетич (Сербия-Болгария-Республика Македония-Россия-Франция)
- Лаура Королева, «Света», реж. Жанна Исабаева (Казахстан)
Лучшая музыкальное решение
- Евгений Гальперин, «Нелюбовь», реж. Андрей Звягинцев (Россия)
- Александр Хант, Константин Шевелев, «Как Витька Чеснок вез Леху Штыря в дом инвалидов», реж. Александр Хант (Россия)
- Яцашек, «Ноябрь», реж. Райнер Сарнет (Эстония-Нидерланды-Польша)
Лучшее художественное оформление
- Ирина Очина, «Мешок без дна», реж. Рустам Хамдамов (Россия)
- Яагуп Роомет, Матис Мэесалу, «Ноябрь», реж. Райнер Сарнет (Эстония-Нидерланды-Польша)
- Арад Сават, «Фокстрот», реж. Самуэль Маоз (Израиль)
Лучшая операторская работа
- Петр Духовской, Тимофей Лобов, «Мешок без дна», реж. Рустам Хамдамов (Россия)
- Михаил Кричман, «Нелюбовь», реж. Андрей Звягинцев (Россия)
- Март Таниэль, «Ноябрь», реж. Райнер Сарнет (Эстония-Нидерланды-Польша)
Лучший сценарий
- Борис Хлебников, Наталья Мещанинова, «Аритмия», реж. Борис Хлебников (Россия)
- Валеска Гризебах, «Вестерн» (Болгария-Германия-Австрия)
- Олег Негин, Андрей Звягинцев, «Нелюбовь», реж. Андрей Звягинцев (Россия)
- Ильдико Эньеди, «О теле и душе», реж. Ильдико Эньеди (Венгрия)
- Кантемир Балагов, Антон Яруш, «Теснота», реж. Кантемир Балагов (Россия)
 Борис Хлебников (Россия)
Борис Хлебников (Россия)Лучший режиссер
- Кантемир Балагов, «Теснота» (Россия)
- Валеска Гризебах, «Вестерн» (Болгария-Германия-Австрия)
- Андрей Звягинцев, «Нелюбовь» (Россия)
- Самуэль Маоз, «Фокстрот» (Израиль)
Лучший игровой полнометражный фильм
- «Вестерн», реж. Валеска Гризебах (Болгария-Германия-Австрия)
- «Нелюбовь», реж. Андрей Звягинцев (Россия)
- «О теле и душе», реж. Ильдико Эньеди (Венгрия)
- «Теснота», реж. Кантемир Балагов (Россия)
- «Фокстрот», реж. Самуэль Маоз (Израиль)
Церемония вручения первой международной кинематографической премии «Восток — Запад. Золотая арка» состоится в Москве 14 апреля в Центре оперного пения имени Галины Вишневской.
За неделю до вручения премии в Москве, в партнерстве с Гете-Институтом и «Каро Арт» состоятся специальные показы двух фильмов-номинантов «Золотой арки». 6 и 7 апреля в кинотеатре «Каро 8 Атриум» Андрей Плахов представит фильмы «Вестерн» Валески Гризебах (номинации на лучший фильм, лучшую режиссуру, лучший сценарий и лучшую мужскую роль) и «Ноябрь» Райнера Сарнета (номинации на лучшую операторскую работу, лучшую работу художника и лучшее музыкальное решение).
Понравился материал? Помоги сайту!
Подписывайтесь на наши обновления
Еженедельная рассылка COLTA.RU о самом интересном за 7 дней
Лента наших текущих обновлений в Яндекс. Дзен
Дзен
RSS-поток новостей COLTA.RU
При поддержке Немецкого культурного центра им. Гете, Фонда имени Генриха Бёлля, фонда Михаила Прохорова и других партнеров.
«В тесноте – да не в обиде». Как Воронеж принял абсолютный чемпионат России по борьбе. Последние свежие новости Воронежа и области
Чемпионат России по греко-римской и вольной борьбе в абсолютной весовой категории состоялся в Воронеже в субботу, 25 ноября. В соревнованиях, которые прошли в спорткомплексе Дворца творчества детей и молодежи, приняли участие около 80 спортсменов.
«Надеюсь, “абсолютка” всколыхнет былые традиции»
Турниру присвоили имя серебряного призера чемпионата Европы, четырехкратного чемпиона СССР по классической борьбе Василия Меркулова. Он наблюдал за схватками вместе с трехкратным чемпионом Олимпийских игр по греко-римской борьбе, депутатом Госдумы Александром Карелиным, трехкратным олимпийским чемпионом по вольной борьбе Артуром Таймазовым и губернатором Воронежской области Алексеем Гордеевым.
Глава региона поприветствовал собравшихся.
– Очень почетно, что чемпионат России в абсолютной категории проходит у нас. Хочу извиниться, что у нас такой маленький зал. Но, как говорят, в тесноте – да не в обиде. Может, сегодняшние соревнования подтолкнут нас, чтобы построить современный центр спортивной борьбы. Особенную гордость вызывает то, что турнир назван именем выдающегося воронежского спортсмена и тренера Василия Прокофьевича Меркулова. Я думаю, что соревнования «разбудят» наших мальчишек, будущих мужчин, и у нас появятся олимпийские чемпионы. Ведь борьба, классическое единоборство – это самый красивый мужественный вид спорта. Сегодня подобралась удивительная команда участников. Многие являются победителями международных соревнований, есть призеры Олимпийских игр, – отметил Алексей Гордеев.
Губернатор возглавил оргкомитет турнира, согласившись с предложением Александра Карелина. В ходе церемонии открытия турнира Алексей Гордеев вручил Василию Меркулову знак отличия «За заслуги перед Воронежской областью». А Александр Карелин был удостоен знака правительства региона «Благодарность от земли Воронежской».
В ходе церемонии открытия турнира Алексей Гордеев вручил Василию Меркулову знак отличия «За заслуги перед Воронежской областью». А Александр Карелин был удостоен знака правительства региона «Благодарность от земли Воронежской».
Василий Меркулов признал, что с трудом подбирает слова из-за волнения. Он подчеркнул, что впервые в его родной город съехались сильнейшие борцы страны.
– Сегодня лучшие борцы страны выясняют, кто из них сильнейший, в Воронеже. И я благодарен Алексею Гордееву за высокую оценку моих далеко не всегда последовательных инициатив. Можно вспомнить достижения Меркулова и его последователей и сразу понять: Воронеж – территория борьбы. И придать этим традициям более звонкое звучание помогает наш чемпионат. Компактный, конечно, по проведению, но очень наглядный. Надеюсь, что «абсолютка» позволит всколыхнуть былые традиции и сделать так, чтобы у нас появились самые бесспорно сильные тяжеловесы в мире, – заявил Александр Карелин.
Спортсмены и тренеры отмечали, что для проведения абсолютного чемпионата России на регулярной основе в Воронеже необходимо улучшать спортивную инфраструктуру региона. Зал спорткомплекса не мог вместить всех желающих увидеть схватки лучших борцов страны из-за небольших трибун, а сооружение не имеет гардероба – воронежцам приходилось наблюдать за турниром, стоя вокруг ковров с зимними куртками в руках.
«Воронежские борцы выступили на “четыре с минусом”»
В турнире приняли участие спортсмены полутяжелого и тяжелого веса: 98 и 130 кг в греко-римской борьбе и 97 и 125 кг – в вольной борьбе. Воронежскую область на чемпионате представляли Никита Шкарин, Игорь Грошев, Иван Рощупкин, Константин Юрканов (греко-римская борьба), а также Алексей Алаев, Евгений Кинишов и Степан Баев (вольная борьба). Баев стал бронзовым призером соревнований в весовой категории 97 кг.
– Следил за выступлениями ребят, оцениваю их на «четыре с минусом». Могли побороться за большее. Но это объективно пока наш уровень на фоне звезд всероссийского уровня, которые сюда съехались. Конкуренция была высочайшая. Порадовал Баев, у него хороший потенциал. И это при том, что по меркам тяжеловесов у него детский возраст – 20 лет. В эти годы они только начинают формироваться. Так что я настроен оптимистично, – сказал старший тренер Воронежской области по вольной борьбе Геннадий Холодков.
Абсолютный чемпионат России прошел в Воронеже в пятый раз. Однако впервые, как отметили специалисты, он прошел на столь высоком уровне с сильнейшим составом участников.
– Здорово, что он прошел в невиданном до этого момента формате – одновременно борются спортсмены вольного и греко-римского стиля, все в одном зале. Состав участников очень сильный. Гораздо лучше, чем было раньше. Все это состоялось благодаря работе федерации спортивной борьбы России, организационному комитету, который возглавил губернатор. До всего сообщества борцов получилось донести, что в Воронеже состоится реально серьезный турнир, на который обязательно стоит ехать. И увеличение призового фонда тоже сказалось, это абсолютно нормально. Были привлечены внебюджетные средства, что очень важно. И федерация борьбы Воронежской области активно работает в этом направлении, успехи есть. В сборной страны есть воронежец, будут и другие «звездочки». Но загадывать не стоит. Недаром слово «борьба» – женского рода. Борьба очень изменчива. И сегодняшний турнир положительно скажется на развитии нашего вида спорта в регионе. Это ведь популяризация. У воронежцев появилась возможность увидеть вживую Махова, призера Олимпиады. Разве когда-нибудь у нас такое было?А значит, улучшение качества отбора перспективных спортсменов, – признал президент федерации спортивной борьбы Воронежской области Андрей Зенин.
Результаты
Спортивная борьба. Абсолютный чемпионат России. Мужчины. Воронеж
Абсолютный чемпионат России. Мужчины. Воронеж
Вольная борьба
До 97 кг
1. Батрадз Газзаев.
2. Магомед Ханапиев.
3. Степан Баев и Сергей Шмелев.
До 125 кг
1. Билял Махов.
2. Алан Хугаев.
3. Зелимхан Хизриев и Хасан Хубаев.
Абсолютная весовая категория
1. Билял Махов.
2. Батрадз Газзаев.
Греко-римская борьба
До 98 кг
1. Кантемир Магомедов.
2. Никита Мельников.
3. Ахмед Мусаев и Данила Сотников.
До 130 кг
1. Алексей Гришин.
2. Нохчо Лабазанов.
3. Зураби Гедехаури и Лом-али Акаев.
Абсолютная весовая категория
1. Кантемир Магомедов.
2. Алексей Гришин.
Фото — Андрей Архипов
prev next
To view this video please enable JavaScript, and consider upgrading to a web browser that supports HTML5 video
В Воронеже за титул чемпиона России сразились лучшие борцы страны
Заметили ошибку? Выделите ее мышью и нажмите Ctrl+Enter
В тесноте — Справочник химика 21
Оценка тесноты нелинейной связи. Если считать, что уравнение регрессии найдено с достаточной точностью, то остаточная дисперсия обусловлена только наличием дисперсии воспроизводимости, т. е. [c.145]Нетрудно себе представить тесноту и метеорологические условия в таком помещении. Несмотря на наличие принудительной вентиляции, температура воздуха в этом помещении постоянно держится в пределах 50—60 °С, а температура оголенной поверхности арматуры — значительно выше 100 °С. [c.240]
Применяемый на промышленных предприятиях ток для освещения вызывает смертельное поражение.
 Безопасным считается напряжение не выше 36 В, а нри работе в особо опасных условиях (когда возможность поражения увеличивается теснотой) в сырых помещениях, внутри резервуаров и аппаратов безопасным напряжением считается 12 В. [c.419]
Безопасным считается напряжение не выше 36 В, а нри работе в особо опасных условиях (когда возможность поражения увеличивается теснотой) в сырых помещениях, внутри резервуаров и аппаратов безопасным напряжением считается 12 В. [c.419]Тесноту связи между случайными величинами характеризуют корреляционным отношением [c.23]
Как показали проведенные нами статистические исследования (анализ тесноты связи и уравнений регрессий) по изучению влияния условий залегания на состав нефтей разных генотипов, масштабы преобразования нефтей при процессах окисления, физического выветривания, катагенеза и миграции неодинаковы. Нефти разных генотипов могут существенно изменяться при одинаковых термобарических условиях. В связи с этим на одних и тех же глубинах нефти разных генотипов могут иметь разный состав. Последнее обстоятельство очень важно при прогнозировании типа [c.10]
Безопасным напряжением считается напряжение не выше 36 В, а при работе в особо опасных условиях, когда возможность поражения увеличивается теснотой, в сырых помеш,ениях, внутри резервуаров и аппаратов безопасным напряжением считается 12 В. [c.274]
Слово л ктoн происходит от названия лактоновой кислоты, молекула которой содержит и карбоксильную, и гидроксильную группы. Тем не менее лактонов лактоновая кислота не образует, потому что эти группы расположены в молекуле слишком близко друг к другу и не могут сконденсироваться. (В виде примера представьте себе, что вы едете в битком набитом вагоне метро и из-за тесноты не можете пожать руку человеку, стоящему рядом с вами.) Две молекулы лактоновой кислоты могут конденсироваться, образуя так называемый лактид, но об этом соединении мы здесь говорить не будем. [c.189]
Модели показывают, что в средних циклах нет углового (байеровского) напряжения и построены они из нечетных конформаций, т. е. питцеровского напряжения в них тоже нет. Причина повышенной энергии этих циклов — внутримолекулярная теснота , приводящая к тому, что несвязанные атомы вынуждены располагаться на расстояниях, меньших, чем суммы их ван-дер-ваальсовых радиусов. Такой тип напряжения мы назвали ранее (см. стр. 317) прелоговским. В расчете на одну СНз-группу напряжение составляет в циклооктане 5,0 кДж/моль, в циклононане 5,9 кДж/моль, в циклодекане 5,0 кДж/моль, в циклоундекане 4,2 кДж/моль. В циклододекане эта величина падает до 1,3 кДж/моль на группу СН2, и это ясно указывает, что данный цикл уже не [c.369]
Такой тип напряжения мы назвали ранее (см. стр. 317) прелоговским. В расчете на одну СНз-группу напряжение составляет в циклооктане 5,0 кДж/моль, в циклононане 5,9 кДж/моль, в циклодекане 5,0 кДж/моль, в циклоундекане 4,2 кДж/моль. В циклододекане эта величина падает до 1,3 кДж/моль на группу СН2, и это ясно указывает, что данный цикл уже не [c.369]
Если зависимость между значениями случайных функций и t) и у (коэффициент корреляции между значениями случайной функции уже не может служить достаточно хорошим критерием для измерения тесноты связи между ними. Поэтому для характеристики связи между илу используются [c.438]
Выделение режимных координат может, например, осуществляться ранжировкой промежуточных координат по коэффициенту корреляции, характеризующих их связь с критерием, и выбором тех, которые обладают наибольшей теснотой связи. При этом выбор режимных координат будет содержать элемент субъективности. [c.34]
Установить связь можно с помощью группировки, но определить тесноту связи можно только путем составления уравнения корреляции и определения коэффициента (г) или индекса (р) корреляции. Уравнения корреляции являются по существу оттисками теоретической линии регрессии, в которой сумма квадратов отклонений фактических значений варьирующего признака от вычисленных по уравнению была бы наименьшей из всех возможных (на основе способа наименьших квадратов). [c.143]
Он характеризует тесноту линейной связи между случайными величинами X и у. л , [c.56]
Наиболее распространенной формой выражения тесноты линейной связи между двумя случайными величинами является коэффициент корреляции. Его определяют отношением [c.56]
Оценка тесноты связи между интересующими нас параметрами х и у может быть получена в результате определения нормированной корреляционной функции связи. [c.8]
Для прогнозирования коэффициента продуктивности по рассмотренным показателям учитывалась информативность каждого геофизического метода по месторождению. При недостаточном количестве данных для одного месторождения зависимости строили по группе месторождений со сходными геолого-физическими свойствами коллекторов и нефтей. Кроме того, для расчетов выбирали уравнения, характеризующиеся лучшей теснотой связи по коэффициенту корреляции и минимальной погрешностью получаемого результата. [c.86]
При недостаточном количестве данных для одного месторождения зависимости строили по группе месторождений со сходными геолого-физическими свойствами коллекторов и нефтей. Кроме того, для расчетов выбирали уравнения, характеризующиеся лучшей теснотой связи по коэффициенту корреляции и минимальной погрешностью получаемого результата. [c.86]
На втором этапе по геофизическому показателю, характеризующемуся лучшей теснотой связи с коэффициентом продуктивности, и эффективной нефтенасыщенной толщине (в интервалах перфорации исследуемых пластов) рассчитывали зависимости их от коэффициента продуктивности. [c.86]
Коэффициент корреляции г. Для описания тесноты связи (корреляции) параметров х и у в линейном регрессионном анализе используют коэффициент корреляции г, выборочное значение которого вычисляют по формуле [c.40]
Ожог от кислот чаще всего может быть получен при переливании и прикосновении пальцами к кислоте, растекающейся от горлышка склянки по наружной ее части. При практических работах, проводимых на приспособленных партах, вследствие тесноты, а также неустойчивости небольших склянок нередки случаи разливания кислот. При этом возможен ожог рук. Для [c.43]
Само собой разумеется, что для молекул более сложных, чем этан, оказывается возможным существование большого числа конформеров, из которых наиболее устойчивые будут отбираться по тому же принципу — уменьшению внутримолекулярного взаимодействия атомов, что ведет к снижению общей энергии молекулы. Мы имеем при этом в виду невыгодные пространственные взаимодействия (иначе говоря, излишнюю тесноту расположения), в то время как сближение разноименных зарядов будет, наоборот, стабилизировать соответствующую конформацию. [c.71]
В зависимости от того же малого межъядерного расстояния и тесноты пространства, в котором двигаются в областях перекрывания многочисленные связевые электроны основного состояния N2, находится и значительная корреляция этих движений, и соответствующее увеличение энергии молекулярной связи. [c.235]
[c.235]
КО, как следует из выражения (172), изменяется от О до 1 и характеризует тесноту корреляционной связи, в данном случае между колебаниями дебитов различных скважин. Квадрат корреляционного отношения показывает долю от общей дисперсии выходной переменной и объясняется дисперсией набора входных переменных. Следовательно, чем выше КО, тем выше теснота корреляционной связи между выходными и входными параметрами. [c.222]
Последовательно принимается дебит каждой скважины в выбранных группах за выходной параметр, а остальных — за входные, т.е. одна из скважин принимается за реагирующую , а другие — за возмущающие , производится определение тесноты связи между колебаниями дебитов по каждой группе скважин. Расчеты КО вьшолняются с применением ЕС ЭВМ по программе метода группового учета аргументов [5]. [c.223]
СНг- и СНз-групп, изотопному составу углерода этой фракции, соотношению бензольных, нафталиновых и фенантреновых У В, по различиям в индивидуальном составе полициклических аренов [1]. Параметры, характеризующие структуру УВ, как показали данные корреляционнорегрессионного анализа [5, 11], имеют наименьшую тесноту связи с геологическими условиями залегания, т. е. они меньше всего подвержены влиянию внешних факторов. Изучение УВ высококипящих фракций нефтей позволило установить сходство между нефтями и ОВ пород в пределах одного и того же стратиграфического комплекса. Так, в ряде регионов (Предкавказье, Волго-Урал) наблюдалась близость между нефтями и ОВ материнских пород по таким показателям, как число нафтеновых циклов в молекуле парафино-нафтеновой фракции, индивидуальный состав полициклических ароматических УВ, и. с. у. нефтей и фракций. Отмечается однонаправленность изменений этих параметров по разрезу как в нефтях, так и в ОВ, что свидетельствует об их унаследованности нефтями от ОВ материнских пород. [c.10]
Состав и свойства нефтей различных генотипов, приуроченных к отложениям разного возраста, неодинаково изменяются в зависимости от современных геологических условий. Корреляционно-регрессионный анализ показал, что теснота связей и набор коррелируемых параметров неодинаковы. Так, например, в Предкавказье состав нефтей в верхнемеловых отложениях практически не коррелируется с условиями залегания, для юрских нефтей получены значимые коэффициенты между глубиной и углеводородным составом бензиновых фракций. Состав нефтей в нижнемеловых отложениях тесно связан с глубиной залегания, минерализацией и сульфатностью вод. На плиоценовые нефти существенно влияют глубина залегания, температура недр и минерализация пластовых вод. Состав нефтей в олигоценовых и эоценовых отложениях коррелируется (но слабее, чем в плиоценовых) с глубиной, температурой, минерализацией вод, а для миоценовых нефтей, состав которых более тесно связан с условиями залегания, о чем свидетельствуют более высокие коэффициен- [c.147]
Корреляционно-регрессионный анализ показал, что теснота связей и набор коррелируемых параметров неодинаковы. Так, например, в Предкавказье состав нефтей в верхнемеловых отложениях практически не коррелируется с условиями залегания, для юрских нефтей получены значимые коэффициенты между глубиной и углеводородным составом бензиновых фракций. Состав нефтей в нижнемеловых отложениях тесно связан с глубиной залегания, минерализацией и сульфатностью вод. На плиоценовые нефти существенно влияют глубина залегания, температура недр и минерализация пластовых вод. Состав нефтей в олигоценовых и эоценовых отложениях коррелируется (но слабее, чем в плиоценовых) с глубиной, температурой, минерализацией вод, а для миоценовых нефтей, состав которых более тесно связан с условиями залегания, о чем свидетельствуют более высокие коэффициен- [c.147]
Гипергенные и катагенные изменения нефтей определяются геологическими условиями их залегания. Уравнения регрессии, отражающие зависимости между параметрами, неодинаковы по набору параметров состава нефти и по тесноте связи с разными геологическими показателями для разных циклов. В зависимости от времени нахождения нефтей в зоне гипергенеза или в зоне катагенеза с температурой выше той, действие которой испытали материнские породы в палеотемпературной зоне активной генерации и эмиграции масштаб вторичных изменений нефтей будут разный. Отсюда вытекает необходимость для правильного прогнозирования состава нефти изучения ее палеотемпературной истории и количественно выраженной тесноты связи с геологическими условиями залегания. [c.183]
В рабочем столе или шкафу следует держать только самую необходимую, постоянно используемую посуду. Минимальный запас посуды необходим, однако он должен храниться отдельно. Если иосуда не имеет своего постоянного места, хранится неаккуратно, в тесноте, она неизбежно разобьется. [c.16]
Стохастическая связь. Между случайными величинами обычно существует такая связь, при которой с изменением одной величины меняется распределение другой. Такая связь называется стохастической. Изменение случайной величины У, соответствующее изменению величины X, разбивается при этом на две компоненты стохастическую (связанную с зависимостью К от Л») и случайную. Если первая компонента отсутствует, то величины У и X независимы. Если отсутствует вторая компонента, У X связаны функциональной зависимостью. При наличии обеих компонент соотнощение между ними определяет силу (тесноту) связи. В обще.м виде задача выявления и оценки силы стохастической связи в математической статистике не рещена. Существуют показатели, оценивающие те или иные стороны стохастической связи. Из них важнейшим является коэффициент корреляции. [c.126]
Такая связь называется стохастической. Изменение случайной величины У, соответствующее изменению величины X, разбивается при этом на две компоненты стохастическую (связанную с зависимостью К от Л») и случайную. Если первая компонента отсутствует, то величины У и X независимы. Если отсутствует вторая компонента, У X связаны функциональной зависимостью. При наличии обеих компонент соотнощение между ними определяет силу (тесноту) связи. В обще.м виде задача выявления и оценки силы стохастической связи в математической статистике не рещена. Существуют показатели, оценивающие те или иные стороны стохастической связи. Из них важнейшим является коэффициент корреляции. [c.126]
Коэффициент корреляции характеризует ие всякую зависимость, а толььо линейную. Линейная вероятностная зависимость случайных вели нн заключается в том, что при возрастании одной случайной величины другая имеет тенденцию возрастать (или убывать) по линейному закону. Коэффициент корреляции характеризует степень тесноты лгшенной зависимости. Если случайные величины X н V связаны точной линейной функциональной зависимостью у= = Ьо- -Ь1Х, то Лс /= 1 причем знак соответствует знаку коэффициента Ьь В общем случае, когда величины X и У связаны произвольной стохастической зависимостью, коэффициент корреляции может иметь значение в пределах [c.127]
Полученное уравнение является окончательным и классе полиномов. Оценим тесноту найденной связи нри помощи корреляцноиного отношения (IV.89) [c.144]
Если 0=1, то существует функциональная записимость между параметрами. Однако при 0 = 0 величины и X нельзя считать независимыми, так как связь между ними, не сказываясь на дисперсиях, может проявить себя в моментах более высокого порядка. И только при нормал1)Ном раснределеинн равенство нулю корреляционного отиошения однозначно свидетельствует. об отсутствии связи между случайными величинами. Корреляционное отношение, как и коэффициент коррелящш в линейной регрессии, характеризует тесноту связи между случайными величинами. Вообще анализ силы связи по 0 называют корреляционным анализом. [c.146]
Вообще анализ силы связи по 0 называют корреляционным анализом. [c.146]
Это значит, то такой ток, применяемый для осве щения, может вызвать смертельное поражение, не говоря уже о наиболее часто применяемом тОке в, 220 В. Поэтому совершенно неправильным являетсй мнение, ЧТО ток, используемый для освешения, безопасен. Такие взгляды существуют даже у электромонтеров, которые, исходя из своего личного опыта, доказывают, что они подвергались действию тока в 127 и 220 В и йе полу чили электрического удара. Те из них, с которыми это было, просто счастливо отделались, потому что в это время сопротивление их кожи случайно оказалось значительно выше 1000 Ом. Статистика показывает, что наибольшее число несчастных случаев происходит именно при напряжениях 127 и 220 В. Анализ несчастных случаев показывает, что смертельные исходы возможны при напряжениях, начиная с 60 В. На основании этого безопасным напряжением согласно действующим правилам считается напряжение не свыще 36 В, а при работе в особо опасных условиях, когда/ возможность поражения увеличивается теснотой, в сырых помещениях, внутри резервуаров и аппаратов безопасным напряжением считается 12 В. [c.221]
Часто химические цехи и установки относятся к числу особо опасных по степени поражения работающих электрическим током вследствие сырости, наличия токопроводящих полов и конструкций, иногда тесноты на рабочем месте, поэтому электросварщики должны работать в диэлектрических перчатках, защитных калошах, в изолирующих шлеме, подлокотниках и наколенниках, стоя на резиновом коврике. Над сварочными устройствами, находящимися на открытом воздухе, делаются иавесы, для того чтобы осадки не могли нарушить действия изолирующих средств. Если этого не сделано, то во время дождя или снегопада электросварочные работы прекращаются. Электросварочная аппаратура обязательно заземляется. [c.249]
В то время они с Одил жили в Зеленой Двери — крохотной, недорогой квартирке на верхнем этаже старинного здания напротив Сент-Джонс-колледжа. Только две их комнаты — гостиная и спальня — более или менее заслуживали этого названия. Остальные же помещения, включая и кухню, которую почти всю занимала ванна, практически не существовали вовсе. Однако, несмотря на тесноту, квартирка выглядела очень уютной, если не сказать кокетливой, благодаря художественному вкусу Одил. Там я впервые почувствовал бодрящую силу английской интеллектуальной жизни, о которой я и не подозревал первые дни, проведенные в викторианской комнате на Джезус-Грин в нескольких сотнях ярдов оттуда. [c.44]
Только две их комнаты — гостиная и спальня — более или менее заслуживали этого названия. Остальные же помещения, включая и кухню, которую почти всю занимала ванна, практически не существовали вовсе. Однако, несмотря на тесноту, квартирка выглядела очень уютной, если не сказать кокетливой, благодаря художественному вкусу Одил. Там я впервые почувствовал бодрящую силу английской интеллектуальной жизни, о которой я и не подозревал первые дни, проведенные в викторианской комнате на Джезус-Грин в нескольких сотнях ярдов оттуда. [c.44]
Экономические явления характеризуются многомерной системой различных факторов. Часто им свойственны такие черты, как случайность и неопределенность, стохастический (вероятностный) характер связи между явлениями. В этом случае для изу- ения тесноты связи и взаимосвязи показателей и факторов используют корреляционный и регрессионный анализ. С помощью этих методов можно количественно оценить степень влияния того или иного фактора. Например, только используя корреляционный анализ, можно оценить степень влияния квалификационного состава рабочих, стажа их работы на производительность труда, объем чистой продукции или себестоимость. Бесспорно, что производительность труда зависит от стажа работы, но этот фактор не является определяющи.м, так как на производительность труда влияют уровень техники, организации производства и труда, а также другие факторы, т. е. связь в данном случае не функ-ипональная, а корреляционная. [c.142]
Эти временные мероприятия затянулись на целых десять лет, когда работники в условиях недопустимой тесноты, загазованности низких помещений обеспечивали потребность оборонной техники в своей продукции. Все эти годы производство силицированного графита вели молодые инженеры, ставшие постепенно опытными специалистами. Это Фридрих Геннадьевич Кудрецкий, Анатолий Михайлович Зыков, молодой инженер Седов. А на переделе механической обработки силицированного графита хорошо зарекомендовал себя И. И. Савельев. [c.100]
И. Савельев. [c.100]
Такое агрегирование происходит вследствие тесноты , когда частицы даже независимо от броуновского движения оказываются на расстояниях, при которых они принуждены скатываться в первичную потенциальную яму. В этом случае нарушение агрегативной устойчивости пройсхс/дит по механизму, отличному от коагуляции электролитами. [c.296]
Метро уходит в отпуск | Статьи
Завсегдатаи московского метро жалуются: лето, кризис, а в вагонах по-прежнему теснота. Наверное, подземные перевозчики решили сэкономить на удобстве пассажиров и сократили количество поездов. Руководство метрополитена уверяет: в данный момент завершается переход на летний график, который обычно вводится с мая по сентябрь. Нынешнее лето, говорят они, в этом смысле ничем не отличается от предыдущих.
Минимальный интервал, технологически допустимый для метро, — 90 секунд. Среднестатистический никто не считал, но если «на глазок», то это примерно 2-2,5 минуты. На летний период интервал между поездами в среднем увеличен на 15 секунд. Например, одной из первых «под сокращение» попала Таганско-Краснопресненская линия. В непиковое время поезда стали приходить на ее станции не через каждые 2 минуты 10 секунд, а через 2 минуты 30 секунд.
— Непиковое время, — пояснили «Известиям» в пресс-службе метрополитена, — это три промежутка: с открытия метро до 7.00, с 11.00 до 17.00 и с 21.00 до закрытия.
В часы пик график не меняется. И хотя рейсов в течение суток в итоге становится меньше, сотрудникам метрополитена от этого не легче. Ведь лето — период отпусков. А в отдельных случаях работы даже прибавляется. Например, на короткой Каховской линии (Каховская-Варшавская-Каширская) много времени занимает, как выражаются в метро, оборот (разворот) состава в тоннеле-тупике для поездки в обратном направлении. Работая в обычном графике, один машинист доводил состав до конечной станции, а там в кабину на другом конце поезда садился второй. После чего первый вел состав к обороту. Там второй переключал управление на себя и возвращал поезд на перрон. Летом оставляют только одного машиниста, которому приходится бегать вдоль поезда. Что, конечно, тоже удлиняет срок ожидания пассажиров.
Там второй переключал управление на себя и возвращал поезд на перрон. Летом оставляют только одного машиниста, которому приходится бегать вдоль поезда. Что, конечно, тоже удлиняет срок ожидания пассажиров.
Зато в поездах, следующих по Калининской линии (Новогиреево-Третьяковская), значительно прибавится свободных мест. Пассажиропоток падает, но именно сейчас здесь увеличивают число вагонов. В каждом составе их теперь не 7, а 8. Схему движения восьмивагонных поездов отрабатывали с января: передвигали место остановки головного вагона, перенастраивали автоматику.
Это третья ветка метро, где ходят 8-вагонные поезда. Впервые в 2006 году восьмые вагоны прицепили к составам на Серпуховско-Тимирязевской ветке, в прошлом году — на Люблинской.
Самой комфортабельной линией обещают сделать Кольцевую. Скоро закончится обкатка поезда «Русич» нового образца. У старого «Русича» 4 двери с каждой стороны, у нового — 6. Следовательно, посадка и высадка будут происходить быстрее.
РАСТЯГИВАНИЕ И ГИБКОСТЬ — Физиология растяжения
Перейти к предыдущей, следующей главе.
Цель этой главы — познакомить вас с некоторыми из основных физиологические концепции, которые вступают в действие при растяжении мышцы. Сначала будут представлены концепции с общим обзором, а затем (для желающих узнать кровавые подробности) будет обсуждаться в дальнейшие детали. Если вас не очень интересует этот аспект растяжка, вы можете пропустить эту главу.Другие разделы будут ссылаться на важные концепции из этой главы, и вы можете легко найти их основа «необходимости знать».
Вместе мышцы и кости составляют то, что называется костно-мышечная система тела. Кости обеспечивают осанку и
структурная поддержка тела и мускулов обеспечивают телу
способность двигаться (сокращаясь и тем самым создавая напряжение). В
костно-мышечная система также обеспечивает защиту организм внутреннего
органы. Чтобы выполнять свою функцию, кости должны быть соединены вместе.
чем-то. Точка, где кости соединяются друг с другом, называется сустав , и это соединение в основном связок (вместе с помощью мышц). Мышцы прикрепляются к кости с помощью сухожилий . Кости, сухожилия и связки не обладают способностью
(как мышцы), чтобы ваше тело двигалось. Мышцы очень уникальны в этом
уважать.
Точка, где кости соединяются друг с другом, называется сустав , и это соединение в основном связок (вместе с помощью мышц). Мышцы прикрепляются к кости с помощью сухожилий . Кости, сухожилия и связки не обладают способностью
(как мышцы), чтобы ваше тело двигалось. Мышцы очень уникальны в этом
уважать.
Мышцы различаются по форме и размеру и служат разным целям. Большинство крупных мышц, таких как подколенные сухожилия и квадрицепсы, контролируют движения.Другие мышцы, такие как сердце и мышцы внутреннего уха, выполняют другие функции. Однако на микроскопическом уровне все мышцы разделяют та же основная структура.
На самом высоком уровне (целая) мышца состоит из множества нитей ткань называется пучков . Это мышечные волокна, которые мы посмотрите, когда мы режем красное мясо или птицу. Каждый пучок состоит из пучков , которые представляют собой пучки мышечных волокон . Мышца волокна, в свою очередь, состоят из десятков тысяч нитевидных миофибриллы , которые могут сокращаться, расслабляться и удлиняться (удлиняться).Миофибриллы (в свою очередь) состоят из миллионов полосок, уложенных из конца в конец набрал саркомеров . Каждый саркомер сделан из перекрывающиеся толстые и тонкие филаменты называются миофиламентами . В толстые и тонкие миофиламенты состоят из сократительных белков , в первую очередь актин и миозин.
Как сокращаются мышцы
То, как работают все эти различные уровни мышц, как
следующим образом: Нервы соединяют позвоночник с мышцами.Место, где
соединение нерва и мышцы называется нервно-мышечным соединением .
Когда электрический сигнал проходит через нервно-мышечное соединение, он
передается глубоко внутрь мышечных волокон. Внутри мышечных волокон
сигнал стимулирует поток кальция, который вызывает густой и
тонкие миофиламенты скользят друг по другу. Когда это происходит, это
заставляет саркомер укорачиваться, что создает силу. Когда миллиарды
саркомеры в мышцах сразу укорачиваются, что приводит к сокращению
всего мышечного волокна.
Когда миллиарды
саркомеры в мышцах сразу укорачиваются, что приводит к сокращению
всего мышечного волокна.
Когда мышечное волокно сокращается, оно сокращается полностью. Нет таких вещь как частично сокращенное мышечное волокно. Мышечные волокна неспособны варьировать интенсивность их сжатия по отношению к нагрузке против которые они действуют. Если это так, то как сила мышечные сокращения различаются по силе от сильного до слабого? Что случается состоит в том, что задействуется больше мышечных волокон, если они необходимы для выполнения работа под рукой. Чем больше мышечных волокон задействовано центральной нервной системы, тем сильнее сила, создаваемая мышечной сокращение.
Быстрые и медленные мышечные волокна
Энергия, которая производит поток кальция в мышечных волокнах, поступает от митохондрии , часть мышечной клетки, которая преобразует глюкозу
(уровень сахара в крови) в энергию. Различные типы мышечных волокон имеют
разное количество митохондрий. Чем больше митохондрий в мышце
волокно, тем больше энергии оно способно произвести. Мышечные волокна
подразделяются на медленных волокон и быстро сокращающихся волокон .
Медленно сокращающиеся волокна (также называемые мышечными волокнами типа 1 ) медленно развиваются.
сокращаются, но они также очень медленно утомляются.Быстро сокращающиеся волокна
очень быстро сжимаются и бывают двух разновидностей: Тип 2А
мышечные волокна , которые утомляются со средней скоростью, и типа 2B
мышечные волокна , которые очень быстро утомляются. Основная причина
медленно сокращающиеся волокна медленно утомляются из-за того, что они содержат больше
митохондрии, чем быстро сокращающиеся волокна, и, следовательно, способны производить больше
энергия. Медленно сокращающиеся волокна также меньше в диаметре, чем быстро сокращающиеся.
волокна и увеличивают капиллярный кровоток вокруг них.Потому что они
имеют меньший диаметр и повышенный кровоток, медленные
волокна способны доставлять больше кислорода и удалять больше отходов
из мышечных волокон (что снижает их «утомляемость»).
Эти три типа мышечных волокон (типы 1, 2A и 2B) содержатся в все мышцы в разном количестве. Мышцы, которые нужно сильно сокращать времени (как и сердце) имеют большее количество Тип 1 (медленный) волокна. Когда мышца впервые начинает сокращаться, это в первую очередь 1-й тип. сначала активируются волокна, затем волокна Типа 2А и Типа 2В активируются (при необходимости) в указанном порядке.Дело в том, что мышечные волокна набранных в этой последовательности — это то, что дает возможность выполнять мозговые команды с такими точно настроенными мышечными ответами. Это также делает волокна типа 2B трудно тренировать, потому что они не активируются пока не будет задействовано большинство волокон Типа 1 и Типа 2А.
HFLTA утверждает, что лучший способ запомнить разница между мышцами с преимущественно медленными волокнами и мышцы с преимущественно быстросокращающимися волокнами следует думать о «белых мясо »и« темное мясо ».Темное мясо темное, потому что в нем больше медленно сокращающихся мышечных волокон и, следовательно, большего количества митохондрий, которые темные. Белое мясо состоит в основном из мышечных волокон, которые большую часть времени отдыхают, но их часто просят кратко приступы интенсивной активности. Эта мышечная ткань может быстро сокращаться, но быстро утомляется и медленно восстанавливается. Белое мясо светлее чем темное мясо, потому что в нем меньше митохондрий.
Расположены вокруг мышцы, а ее волокна составляют соединительных волокон.
ткани .Соединительная ткань состоит из основного вещества и двух
виды клетчатки на белковой основе. Два типа волокна: коллагеновая соединительная ткань и эластичная соединительная ткань .
Коллагеновая соединительная ткань состоит в основном из коллагена (отсюда и ее
название) и обеспечивает прочность на разрыв. Эластичная соединительная ткань состоит
в основном из эластина и (как можно догадаться по названию) обеспечивает
эластичность. Основное вещество называется мукополисахарид и
действует как смазка (позволяя волокнам легко скользить по одному
другой), и в качестве клея (удерживая волокна ткани вместе в
связки).Более эластичная соединительная ткань вокруг сустава,
тем больше диапазон движений в этом суставе. Соединительные ткани
состоящие из сухожилий, связок и фасциальных влагалищ, которые охватывают, или
связать мышцы в отдельные группы. Эти фасциальные оболочки или фасция , названы в соответствии с их расположением в
мышцы:
Основное вещество называется мукополисахарид и
действует как смазка (позволяя волокнам легко скользить по одному
другой), и в качестве клея (удерживая волокна ткани вместе в
связки).Более эластичная соединительная ткань вокруг сустава,
тем больше диапазон движений в этом суставе. Соединительные ткани
состоящие из сухожилий, связок и фасциальных влагалищ, которые охватывают, или
связать мышцы в отдельные группы. Эти фасциальные оболочки или фасция , названы в соответствии с их расположением в
мышцы:
- эндомизий
- Самая внутренняя фасциальная оболочка, охватывающая отдельные мышечные волокна.
- перимизий
- Фасциальная оболочка, которая связывает группы мышечных волокон в отдельные fasciculi (см. раздел «Состав мышц»).
- эпимизий
- Внешняя фасциальная оболочка, связывающая пучки целиком (см. Раздел «Состав мышц»).
Эти соединительные ткани помогают обеспечить эластичность и тонус мышцы.
Когда мышцы заставляют конечность перемещаться по диапазону движения сустава, они обычно действуют в следующих сотрудничающих группах:- агонистов
- Эти мышцы вызывают движение. Они создают нормальный диапазон движения в суставе сокращением.Агонисты также называют первичных движителей , поскольку они являются мышцами, которые в первую очередь отвечает за создание движения.
- антагонисты
- Эти мышцы действуют противоположно движению, производимому агонисты и несут ответственность за возвращение конечности в исходное позиция.
- синергистов
- Эти мышцы выполняют или помогают в выполнении одного и того же набора суставов.
движение как агонисты. Синергистов иногда называют нейтрализаторов потому что они помогают нейтрализовать или нейтрализовать лишние
движение от агонистов, чтобы убедиться, что генерируемая сила работает
в желаемой плоскости движения.
- фиксаторы
- Эти мышцы обеспечивают необходимую поддержку, чтобы помочь удерживать остальная часть тела на месте во время движения. Фиксаторы также Стабилизаторы иногда называют .

Например, когда вы сгибаете колено, подколенное сухожилие сокращается и, в некоторой степени, то же самое касается икроножной мышцы и нижней части ягодиц. Между тем, квадрицепсы заторможены (расслаблены и растянуты. несколько), чтобы не сопротивляться сгибанию (см. раздел «Взаимное торможение»).В этом примере подколенное сухожилие служит агонистом или первичный двигатель; квадрицепс служит антагонистом; и теленок и нижние ягодицы служат синергистами. Агонисты и антагонисты обычно располагается на противоположных сторонах пораженного сустава (например, ваш подколенные сухожилия и квадрицепсы или ваши трицепсы и бицепсы), в то время как синергисты обычно располагаются на той же стороне сустава рядом с агонистами. Более крупные мышцы часто призывают своих более мелких соседей действовать как синергисты.
Ниже приводится список наиболее часто используемых мышц-агонистов / антагонистов. пары:
- грудные / широчайшие мышцы спины (грудные и широчайшие)
- передние дельтоиды / задние дельты (переднее и заднее плечо)
- трапеции / дельты (трапеции и дельты)
- брюшной пресс / выпрямители позвоночника (пресс и поясница)
- левый и правый внешние косые (стороны)
- квадрицепсы / подколенные сухожилия (квадрицепсы и бедра)
- голени / икры
- бицепс / трицепс
- сгибатели / разгибатели предплечья
Сокращение мышцы не обязательно означает, что мышца укорачивается; это только означает, что возникло напряжение.Мышцы могут заключить договор следующими способами:
- изометрическое сжатие
- Это сокращение, при котором не происходит никакого движения, потому что нагрузка
на мышцу превышает напряжение, создаваемое сокращающейся мышцей. Это происходит, когда мышца пытается толкать или тянуть неподвижный объект.
- изотоническое сокращение
- Это сокращение, в котором происходит движение и , потому что
напряжение, создаваемое сокращающейся мышцей, превышает нагрузку на
мышца.Это происходит, когда вы используете свои мышцы для успешного толчка или
тянуть объект.
Изотонические сокращения делятся на два типа:
- концентрическое сжатие
- Это сокращение, при котором мышца уменьшается в длину (укорачивается). против встречного груза, например, поднимая тяжесть.
- эксцентрическое сжатие
- Это сокращение, при котором мышца увеличивается в длину. (удлиняется) при сопротивлении нагрузке, например, при нажатии на что-либо.
Во время концентрического сокращения сокращающиеся мышцы служат агонистами и, следовательно, выполняют всю работу. Во время эксцентрическое сокращение, мышцы, которые удлиняются, служат агонисты (и делают всю работу). См. Раздел «Взаимодействие групп мышц».
 Это происходит, когда мышца пытается толкать или тянуть неподвижный объект.
Это происходит, когда мышца пытается толкать или тянуть неподвижный объект. Растяжение мышечного волокна начинается с саркомера.
(см. раздел «Состав мышц»), основная единица сокращения в
мышечное волокно. По мере того как саркомер сжимается, область перекрытия между
толстые и тонкие миофиламенты увеличиваются.По мере того, как он растягивается, эта область
перекрытия уменьшается, позволяя мышечным волокнам удлиняться. Однажды
мышечное волокно находится на максимальной длине покоя (все саркомеры
полностью растянуты), дополнительное растяжение накладывает силу на окружающие
соединительная ткань (см. раздел «Соединительная ткань»). По мере увеличения напряжения
волокна коллагена в соединительной ткани выравниваются вдоль
та же силовая линия, что и напряжение. Следовательно, когда вы растягиваетесь, мышца
волокно вытягивается саркомером на всю длину саркомера, а затем
соединительная ткань принимает оставшуюся слабину.Когда это происходит, это
помогает перестроить любые неорганизованные волокна в направлении
напряжение. Эта перестройка помогает восстановить поврежденную ткань.
вернуться к здоровью.
Эта перестройка помогает восстановить поврежденную ткань.
вернуться к здоровью.
Когда мышца растягивается, некоторые из ее волокон удлиняются, а другие волокна могут оставаться в покое. Текущая длина всей мышцы зависит от количества растянутых волокон (аналогично тому, как общая сила сокращающейся мышцы зависит от количества набираемые волокна сокращаются). Согласно SynerStretch вы следует думать о «маленьких карманах волокон, распределенных по всей растяжение мышечного тела, а другие волокна просто идут на поездка».Чем больше растянуты волокна, тем больше длина развита растянутой мышцей.
Проприорецепторы
Нервные окончания, которые передают всю информацию о опорно-двигательном аппарате Система центральной нервной системы называется проприорецепторов . Проприорецепторы (также называемые механорецепторами ) являются источником всех проприоцепция : восприятие собственного положения тела и движение. Проприорецепторы обнаруживают любые изменения физического смещения (движение или положение) и любые изменения напряжения или силы в пределах тело.Они находятся во всех нервных окончаниях суставов, мышц и сухожилия. Проприорецепторы, связанные с растяжением, расположены в в сухожилиях и в мышечных волокнах.
Существует два вида мышечных волокон: интрафузальных мышечных волокон, и экстрафузальные мышечные волокна . Волокна Extrafusil — это те волокна, которые
содержат миофибриллы (см. раздел «Состав мышц») и обычно
имеется в виду, когда мы говорим о мышечных волокнах. Интрафузальные волокна также называются мышечными веретенами и лежат параллельно экстрафузальным волокнам.Мышечные веретена, или рецепторов растяжения , являются первичными
проприорецепторы в мышце. Еще один проприорецептор, который вступает в игру
при растяжении располагается в сухожилии возле конца мышцы
волокно и называется органом сухожилия Гольджи . Третий тип
проприорецептор, называемый пачинским тельцем , расположен недалеко от
орган сухожилия Гольджи и отвечает за обнаружение изменений в
движение и давление внутри тела.
Когда экстрафузальные волокна мышцы удлиняются, растут и интрафузальные волокна. волокна (мышечные веретена).Мышечное веретено содержит два разных типы волокон (или рецепторов растяжения), чувствительные к изменению длины мышцы и скорости изменения длины мышцы. Когда мышцы сокращается, это создает напряжение в сухожилиях там, где орган сухожилия Гольджи расположен. Орган сухожилия Гольджи чувствителен к изменению напряжения и скорость изменения напряжения.
Рефлекс растяжения
Когда мышца растягивается, растягивается и мышечное веретено. (см. раздел Проприорецепторы).Мышечное веретено фиксирует изменение длины (и как быстро) и посылает сигналы к позвоночнику, которые передают это Информация. Это вызывает рефлекс растяжения (также называемый миотатический рефлекс ), который пытается сопротивляться изменению мышцы длины, заставляя растянутую мышцу сокращаться. Чем внезапнее изменение длины мышцы, тем сильнее будут сокращения мышц (плиометрическая, или «прыжковая» тренировка основана на этом факте). Этот базовый функция мышечного веретена помогает поддерживать мышечный тонус и защитить тело от травм.
Одна из причин проведения растяжки в течение длительного периода времени заключается в том, что когда вы удерживаете мышцу в растянутом положении, мышца веретено приживается (привыкает к новой длине) и сокращает его сигнализация. Постепенно вы можете тренировать свои рецепторы растяжения, чтобы большее удлинение мышц.
Некоторые источники предполагают, что при длительных тренировках растяжка
рефлекс определенных мышц можно контролировать, так что
или отсутствие рефлекторного сокращения в ответ на внезапное растяжение.Пока
этот тип управления дает возможность для максимального
увеличивает гибкость, а также обеспечивает наибольший риск травм
при неправильном использовании. Только непревзойденные профессиональные спортсмены и
считается, что танцоры, достигшие вершины своего вида спорта (или искусства), на самом деле
обладают таким уровнем мышечного контроля.
Компоненты Stretch Reflex
Рефлекс растяжения имеет как динамический, так и статический компоненты. Статический компонент рефлекса растяжения сохраняется до тех пор, пока мышца растягивается.Динамический компонент рефлекса растяжения (который может быть очень мощным) длится всего мгновение и отвечает к первоначальному внезапному увеличению длины мышц. Причина, по которой Рефлекс растяжения состоит из двух компонентов, потому что на самом деле их два виды интрафузионных мышечных волокон: волокон ядерной цепи , которые являются отвечает за статическую составляющую; и ядерных волокон мешка , которые отвечают за динамическую составляющую.
Волокна ядерной цепи длинные и тонкие, при растянуты.Когда эти волокна растягиваются, нервы рефлекса растяжения увеличивать их скорострельность (сигнализацию) по мере того, как их длина постоянно увеличивается. Это статический компонент рефлекса растяжения.
Волокна ядерного мешка выступают посередине, где они наиболее
эластичный. Нервные окончания этих волокон, чувствительные к растяжению, обернуты
вокруг этой средней области, которая быстро удлиняется, когда волокно
растянуты. Внешне-средние области, напротив, действуют так, как будто они
наполнен вязкой жидкостью; они сопротивляются быстрому растяжению, затем постепенно
расширяются при длительном напряжении.Итак, когда требуется быстрое растяжение
у этих волокон поначалу больше всего растягивается середина; тогда, как
внешне-средние части расширяются, средняя может несколько укорачиваться. Так что
нерв, который ощущает растяжение этих волокон, быстро срабатывает вместе с
начало быстрого растяжения, затем замедляется по мере того, как средний участок волокна
разрешено снова сокращаться. Это динамический компонент
рефлекс растяжения: сильный сигнал к сокращению в начале быстрого
увеличение длины мышц, за которым следует немного «выше нормы»
сигнализация, которая постепенно уменьшается по мере того, как скорость изменения мышцы
длина уменьшается.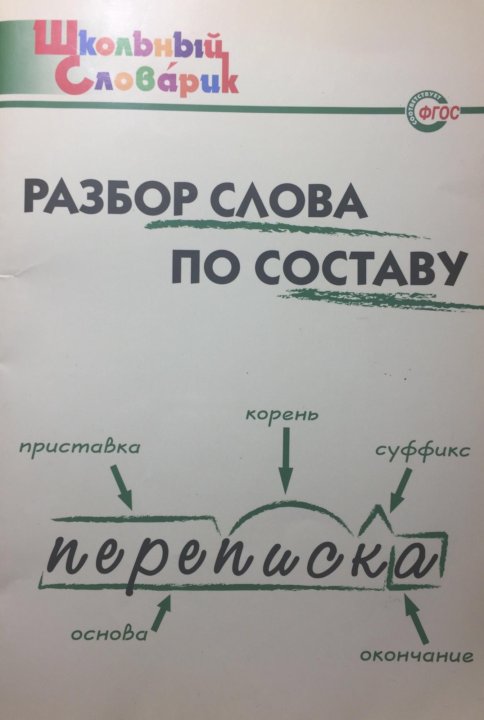
Реакция удлинения
Когда мышцы сокращаются (возможно, из-за рефлекса растяжения), они производят напряжение в точке, где мышца соединяется с сухожилием, где расположен орган сухожилия Гольджи. Орган сухожилия Гольджи регистрирует изменение напряжения, и скорость изменения напряжения, и отправляет сигналы к позвоночнику, чтобы передать эту информацию (см. раздел Проприорецепторы). Когда это напряжение превышает определенный порог, оно запускает реакция удлинения , которая препятствует сокращению мышц и заставляет их расслабиться.Другие названия этого рефлекса — обратный миотатический рефлекс , аутогенное торможение и складной нож reflex . Эта основная функция сухожилия Гольджи орган помогает защитить мышцы, сухожилия и связки от травм. Реакция удлинения возможна только потому, что сигнал Гольджи орган сухожилия спинного мозга достаточно мощный, чтобы преодолеть сигнализация мышечных веретен, сообщающих мышце о сокращении.
Еще одна причина для растяжки в течение длительного периода времени — это чтобы позволить этой реакции удлинения произойти, тем самым помогая растянутому мышцы, чтобы расслабиться.Легче растянуть или удлинить мышцу, когда она не пытается заключить контракт.
Взаимное ингибирование
Когда агонист сокращается, чтобы вызвать желаемое движение, он обычно заставляет антагонистов расслабиться (см. раздел «Взаимодействие групп мышц»). Это явление называется реципрокным торможением , потому что антагонистам запрещено сокращаться. Иногда это называют реципрокная иннервация , но этот термин на самом деле неверный, поскольку он агонисты, которые подавляют (расслабляют) антагонисты.Антагонисты действительно ли , а не , действительно иннервируют (вызывают сокращение) агонистов.
Такое подавление мышц-антагонистов необязательно.
На самом деле может произойти совместное сокращение. Когда вы выполняете приседания, можно
обычно предполагают, что мышцы живота препятствуют сокращению
мышцы поясничного или нижнего отдела спины. В этом конкретном
Однако, например, мышцы спины (выпрямители позвоночника) также сокращаются. Этот
это одна из причин, почему приседания хороши как для укрепления спины, так и для
желудок.
В этом конкретном
Однако, например, мышцы спины (выпрямители позвоночника) также сокращаются. Этот
это одна из причин, почему приседания хороши как для укрепления спины, так и для
желудок.
При растяжении расслабленную мышцу растянуть легче, чем чтобы растянуть сокращающуюся мышцу. Воспользовавшись ситуации, когда происходит реципрокное торможение , можно получить более эффективная растяжка за счет расслабления антагонистов во время растягиваются из-за сокращения агонистов. Вы также хотите расслабиться любые мышцы, которые используются как синергисты мышцы, которую вы пытаетесь растянуть. Например, когда вы растягиваете икры, вы хотите сжать голень. мышцы (антагонисты голени), сгибая стопу.Тем не менее подколенные сухожилия используют икроножные мышцы как синергист, поэтому вы также хотите расслабить подколенные сухожилия, сокращая четырехглавую мышцу (т. е. удерживая ногу прямой).
Перейти к предыдущей, следующей главе.
Статья о герметичности по The Free Dictionary
способность оболочки тела или его отдельных элементов и соединений блокировать обмен газов или жидкостей между средами, разделенными оболочкой. Герметичность — необходимое условие работы многих систем, аппаратов и инструментов.Требуемая герметичность покрытия (допустимая замена) определяется из условий, необходимых для нормального протекания процесса, для которого создается герметичность. Степень герметичности покрытия характеризуется количеством вещества, проходящего через него в единицу времени, и измеряется для жидкостей в литрах в секунду ( · / сек) или граммах в секунду, а для газов и паров: в граммах в секунду или литрах · мм рт. ст. / сек ( л, · мм рт. ст. / сек). Покрытие считается герметичным, если жидкостный или газообмен через него не превышает допустимого уровня.
Герметичность — важное свойство объектов, которое необходимо учитывать при проектировании опломбированных объектов, предназначенных для длительного хранения и использования. Количественные характеристики герметичности объекта определяют его надежность и долговечность. Расчеты производятся для установления вероятного периода времени, в течение которого при определенных условиях использования (колебания давления, температуры, нагрузки и т. Д.) Жидкости или газы могут протекать через все покрытие или его части и нарушать герметичность.
Количественные характеристики герметичности объекта определяют его надежность и долговечность. Расчеты производятся для установления вероятного периода времени, в течение которого при определенных условиях использования (колебания давления, температуры, нагрузки и т. Д.) Жидкости или газы могут протекать через все покрытие или его части и нарушать герметичность.
Обмен газа или жидкости — то есть причина нарушения герметичного состояния — может быть вызван проницаемостью материала покрытия или стыка с неповрежденной структурой, что учитывается при строительстве, или утечками в структура материала или стыков, которые обнаруживаются с помощью вакуумной техники. Осуществляется серийное производство так называемых герметичных отливок для деталей двигателей, турбин, систем водоснабжения и отопления.
Другие объекты, которые должны быть герметичными, включают корпуса самолетов и космических кораблей, подводных лодок, водолазные костюмы и кессонные камеры.Для поддержания сверхвысокого вакуума в полостях термоядерных устройств, ускорителей и космических тренажеров необходима высокая степень герметичности. Нормальная работа таких систем может быть нарушена из-за утечек от 10 -7 до 10 -8 / · мм рт. Еще более строгие требования к герметичности существуют для корпусов электровакуумных и газонаполненных миниатюрных инструментов, в которых давление или состав газовой среды не должны заметно меняться при длительном хранении или в процессе использования.Например, для поддержания рабочего давления в электровакуумном приборе объемом 100 см 3 , не содержащем геттера, утечка в его оболочке не должна превышать 3 x 10 -12 1 · мм рт. срок хранения один год.
Герметичность нефтяных и газовых скважин играет важную роль в процессе бурения залежей с высоким пластовым давлением. Герметизация поверхностей осуществляется при вскрытии нефтяных пластов, когда существует опасность выброса или открытого фонтанирования.Также необходимо, чтобы поверхности были герметичными при разработке нефтяных и газовых месторождений, чтобы предотвратить небольшие утечки минерала между местом бурения и нефтеперерабатывающим заводом. Обслуживание герметичных уплотнений является обязательным во многих технологических процессах в химической, пищевой, фармацевтической и консервной промышленности, когда необходимо поддерживать высокое давление, вакуум или стерильные условия. Необходимо герметизировать промышленное оборудование и инструменты, которые подвергаются воздействию влаги, газов, пыли, грязи, агрессивных химических веществ или инструментов, излучающих радиоактивные лучи.При соблюдении температурно-влажностных и санитарных условий на рабочих местах строятся герметичные производственные здания.
Обслуживание герметичных уплотнений является обязательным во многих технологических процессах в химической, пищевой, фармацевтической и консервной промышленности, когда необходимо поддерживать высокое давление, вакуум или стерильные условия. Необходимо герметизировать промышленное оборудование и инструменты, которые подвергаются воздействию влаги, газов, пыли, грязи, агрессивных химических веществ или инструментов, излучающих радиоактивные лучи.При соблюдении температурно-влажностных и санитарных условий на рабочих местах строятся герметичные производственные здания.
СПИСОК ЛИТЕРАТУРЫ
Смирнов А.И., Сотников А.А. Литейное производство , 1964, вып. 4, стр. 25; нет. 6, стр. 27.Ланис В. А., Левина Л. Е.. Техника вакуумных испытаний , 2-е изд. Москва-Ленинград, 1963.
Большая Советская Энциклопедия, 3-е издание (1970-1979). © 2010 The Gale Group, Inc. Все права защищены.
Границы | Термомеханическая устойчивость и газонепроницаемость стеклокерамических стыков ТОТЭ в системе MgO-BaO / SrO-B2O3-SiO2
Введение
Обработка герметизирующего материала является ключевой проблемой, которая влияет на качество соединения между компонентами батареи ТОТЭ и стеклокерамическим герметиком (Lessing, 2007). Плохая газонепроницаемость отрицательно скажется на характеристиках дымовой трубы и будет способствовать ее ухудшению (Fergus, 2005). Хороший контроль обработки также повлияет на долговременное сопротивление и стабильность соединений в среде сосуществующих окислительной и восстановительной атмосфер клетки (Weil, 2006).Если стекло не обеспечивает хорошую герметизирующую способность, будут зоны легкого доступа для газов на стыках уплотнение / межсоединение и уплотнение / электролит, что будет способствовать деградации границ раздела и, как следствие, ухудшить соединение из-за образования нежелательных продукты коррозии (Haanappel et al., 2005; Rodríguez-López et al., 2016). Кроме того, само уплотнение, а также его соединения с компонентами пакета должны выдерживать различные механические нагрузки, возникающие в результате плоской конфигурации и его модульного характера, включая как сжимающие, так и растягивающие напряжения (Blum et al. , 2011). Еще одним важным качеством является то, что пломбы должны работать как электрические изоляторы между повторяющимися единицами. Как следствие, не только уплотнения должны быть электрическими изоляторами, но также должны хорошо соединяться с материалами трубы (Fergus, 2005). Адгезия стекла коррелирует с его вязкостью при температуре соединения, допустимой для компонентов ТОТЭ. Параллельно межфазные реакции между стеклянным уплотнением и компонентами также регулируются их термодинамическим равновесием при определенной температуре.Таким образом, процесс соединения неизбежно влияет на различные особенности герметика для стекол.
, 2011). Еще одним важным качеством является то, что пломбы должны работать как электрические изоляторы между повторяющимися единицами. Как следствие, не только уплотнения должны быть электрическими изоляторами, но также должны хорошо соединяться с материалами трубы (Fergus, 2005). Адгезия стекла коррелирует с его вязкостью при температуре соединения, допустимой для компонентов ТОТЭ. Параллельно межфазные реакции между стеклянным уплотнением и компонентами также регулируются их термодинамическим равновесием при определенной температуре.Таким образом, процесс соединения неизбежно влияет на различные особенности герметика для стекол.
Измерение механических свойств стыков стали / стеклокерамики затруднено, поскольку на результаты сильно влияют конфигурация испытания и размер образцов, так что компоненты других сил (не только одной объект измерения, т.е. дополнительный нежелательный изгиб) может повлиять на измерение (Nielsen et al., 2008; Smeacetto et al., 2008; Cela Greven, 2014).Хотя в литературе есть много статей, посвященных механическим свойствам, сравнение результатов оказывается затруднительным из-за этой проблемы.
Для сравнения результатов, касающихся прочности соединений, результаты, относящиеся к типу испытания и типу приложенной силы, следует обсуждать отдельно. Как следствие, испытания на растяжение дают значения около 0,5–18,2 МПа (средние значения для различных герметиков), когда они измеряются при комнатной температуре (Rodríguez-López, 2016), причем более высокое значение соответствует стеклокерамике V1649 на основе SrO-La 3 O 3 -B 2 O 3 стеклянная система, разработанная компанией 3M (Timurkutluk et al., 2015). Значения увеличиваются с 16,7 до 31,3 МПа при измерении при комнатной температуре на воздухе и во влажном состоянии H 2 соответственно, как указано для стеклокерамики GC-9 на основе BaO-CaO-La 2 O 3 — ZrO 2 -Al 2 O 3 -B 2 O 3 -SiO 2 стеклянная система (Lin et al. , 2012, 2015). Фактически, предел прочности при растяжении во влажном состоянии H 2 снижается до максимального значения 16,5 МПа при 700 ° C (Stephens et al., 2009) и 11,9 МПа при 800 ° C для стеклокерамики GC-18 и GC-9. , соответственно.Термическое старение при высокой температуре последней стеклокерамики, по-видимому, увеличивает значения тягового сопротивления.
, 2012, 2015). Фактически, предел прочности при растяжении во влажном состоянии H 2 снижается до максимального значения 16,5 МПа при 700 ° C (Stephens et al., 2009) и 11,9 МПа при 800 ° C для стеклокерамики GC-18 и GC-9. , соответственно.Термическое старение при высокой температуре последней стеклокерамики, по-видимому, увеличивает значения тягового сопротивления.
Результаты, соответствующие прочности на сдвиг, полученные при комнатной температуре, дают значения от 0,9 до 47,1 МПа (Stephens et al., 2009). Стеклокерамическая система GC-18 на основе системы BaO-CaO-Al 2 O 3 -SiO 2 -B 2 O 3 (BCASB) дает более высокие значения, измеренные в этом испытании на кручение. Эта стеклокерамика также обеспечивает более высокие значения при высоких температурах (Stephens et al., 2009). Стеклокерамика GC9 демонстрирует увеличение сопротивления сдвигу с увеличением времени термического старения герметика (800 ° C / 1000 ч; Lin et al., 2015).
И, наконец, значения, полученные в результате испытаний на 4-точечный изгиб при комнатной температуре в диапазоне от 22 до 55 МПа (Rodríguez-López, 2016). Стеклокерамика H (система BaO-CaO-V 2 O 5 -ZnO-Al 2 O 3 -B 2 O 3 -SiO 2 ), армированная частицами серебра, обеспечивает здесь более высокое значение (Wei et al., 2015). Тем не менее, стеклокерамика, сокращенно тип B (система BaO-CaO-Al 2 O 3 -SiO 2 ) обеспечивает лучшее значение 30 МПа при соответствующей высокой температуре для этого применения (Zhao, 2013; Мальцбендер и др., 2014). В целом, время термической обработки улучшает также результаты в отношении прочности на изгиб как при комнатной, так и при высокой температуре (Rodríguez-López, 2016).
Целью данной статьи является проиллюстрировать различные исследования, проведенные для улучшения качества некоторых конкретных стеклокерамических соединительных материалов, и продемонстрировать возможность их использования в приложениях SOFC и SOEC. Во-первых, условия герметизации были оптимизированы для двух выбранных композиций в системе MgO-BaO / SrO-B 2 O 3 -SiO 2 . После того, как соединительные материалы оптимизированы, газонепроницаемость измеряется в зависимости от степени кристаллизации стеклокерамики, ее термоциклирования и влияния восстановительной атмосферы на это свойство. Также было изучено электрическое сопротивление при высокой температуре. Впоследствии химическая совместимость стыков сталь / стеклокерамика была оценена посредством анализа поперечных сечений с использованием SEM и EDX.Кроме того, стабильность соединений также изучалась в зависимости от степени кристаллизации, сопротивления в зависимости от термоциклирования и влияния восстановительной атмосферы.
Во-первых, условия герметизации были оптимизированы для двух выбранных композиций в системе MgO-BaO / SrO-B 2 O 3 -SiO 2 . После того, как соединительные материалы оптимизированы, газонепроницаемость измеряется в зависимости от степени кристаллизации стеклокерамики, ее термоциклирования и влияния восстановительной атмосферы на это свойство. Также было изучено электрическое сопротивление при высокой температуре. Впоследствии химическая совместимость стыков сталь / стеклокерамика была оценена посредством анализа поперечных сечений с использованием SEM и EDX.Кроме того, стабильность соединений также изучалась в зависимости от степени кристаллизации, сопротивления в зависимости от термоциклирования и влияния восстановительной атмосферы.
Наконец, механическое сопротивление соединений относительно изгибной нагрузки было охарактеризовано с использованием метода четырехточечного изгиба при комнатной температуре и соответствующих высоких температурах с изменением толщины уплотнения.
Экспериментальная процедура
Подготовка материалов и определение термических и структурных характеристик
В работе исследованы два стеклокерамических состава в системе BaO / SrO-MgO-B 2 O 3 -SiO 2 .Первый состав основан на BaO (27BaO · 18MgO · 7,5B 2 O 3 · 47,5SiO 2 , мол.%), А второй — на основе SrO (27SrO · 18MgO · 10B 2 O 3 · 45SiO 2 мол.%), Названные 7.5B (Ba) и 10B (Sr) соответственно. Подробности плавления этих двух составов стекла ранее сообщались в Rodríguez-López et al. (2017).
Дилатометрические, термические и некоторые механические свойства этих стекол и соответствующей стеклокерамики также можно найти в Rodríguez-López et al.(2017). Стаканы были измельчены в ацетоне с использованием агатовых шариков и сосудов в планетарной мельнице. В испытаниях на герметичность использовались стеклянные порошки с размером частиц D (v, 0,5) = 13 мкм.
Вязкостно-температурные кривые стекол были измерены с использованием метода вращения для низкой вязкости (η <10 4 dPas; температуры выше 950 ° C) и метода изгиба балки для высоких вязкостей (η: 10 13 — 10 9 дПа · с; температура от 650 до 750 ° C).Вязкость стеклокерамических образцов измерялась методом изгиба пучка.
Вязкость при высокой температуре измерялась вискозиметром Хааке с датчиком E1700 в соответствии со стандартом ISO 7884-2 (ISO 7884–2, 1987). Вискозиметр для изгиба луча представлял собой VIS401 (Bahr Thermoanalyse) с трехточечной конфигурацией с расстоянием 40 мм между двумя нижними точками. Образцы, охарактеризованные по этой методике, представляли собой прямоугольные бруски из стекла и стеклокерамики размером ~ 45 × 4 × 3 мм.Стеклокерамические бруски получали одноосным прессованием стеклянных порошков и термообработкой в течение 24 и 100 ч при 800 ° C для состава 7.5B (Ba) и 750 ° C для состава 10B (Sr). Измерения проводились с применением постоянных нагрузок от 10 до 200 г с использованием наклонов нагрева 2 ° C / мин от комнатной температуры до температуры деформации.
Проведеноэкспериментов по дифракции рентгеновских лучей на дифрактометре Rigaku, модель Ultima IV (Rigaku, Tokio, Япония), оборудованном печью и монохроматическим излучением CuKα (λ = 1.5418 Å). Сканирование составляло 10–70 ° (2θ) с шагом 0,02 °. Образцы нагревали со скоростью 2 ° C / мин. Дифрактограммы снимали с разной скоростью 7, 2 или 0,5 град / мин. Температуру стабилизировали в течение 15 мин перед началом сканирования XRD и поддерживали постоянной до конца эксперимента. Для порошков состава 7.5В (Ва) применялась следующая термическая обработка: от комнатной температуры до 1000 ° С в течение 10 мин, охлаждение до 800 ° С и стабилизация в течение 24 ч. Для порошков состава 10В (Sr) применялась термообработка: от комнатной температуры до 850 ° С и стабилизация в течение 10 ч.
Эксперименты по запечатыванию
Печать выполняется двумя разными методами: паста с помощью робота-дозатора и трафаретная печать.
Уплотнительные подложки
Уплотнение находится в прямом контакте с электролитом полуэлемента (электролит / анод) и материалом межсоединения. В качестве соединительного материала использовались два типа стали: Crofer22APU и Crofer22H (ThyssenKrupp AG). Эти стали были специально разработаны для использования в качестве межсоединений для ИТ-ТОТЭ.В зависимости от измеряемых свойств использовались разные толщины и формы. Подложки перед наложением очков очищали этанолом в ультразвуковой ванне в течение 5 мин. Детали состава и некоторые свойства сталей Crofer22H и Crofer22APU можно найти в Data (2010), ThyssenKrupp VDM Gmb (2010). Сталь Crofer22H, разработанная на основе Crofer22APU, демонстрирует лучшую устойчивость к коррозии и меньшее испарение Cr, что снижает загрязнение катода.
Робот-дозатор
Для дозирования роботом с помощью шприца было приготовлено связующее, содержащее 5% этилцеллюлозы, растворенной в α-терпинеоле. Пасту получали смешиванием 82 мас.% Стекла и 18 мас.% Раствора связующего. Пасту вручную смешивали, гомогенизировали и диспергировали с помощью ультразвука, чтобы избежать образования агломератов. Нанесение пасты для стекла осуществлялось с помощью x-y-робота-дозатора, управляемого программой. По этой методике были приготовлены образцы для измерения газонепроницаемости и электрического сопротивления уплотнений.Две стальные квадратные подложки Crofer22APU размером 50 × 50 мм и толщиной 2 мм, одна из них с отверстием диаметром 10 мм, использовались для обеспечения диффузии газа внутри образца (экспозиция в двойной атмосфере). Были нанесены два слоя для нанесения общей массы 1 г между нанесениями слоев; стеклянную пасту сушили в термокамере при 55 ° C. Для поддержания минимальной толщины уплотнения использовались прокладки из диоксида циркония 180 мкм.
Трафаретная печать
Этот метод нанесения герметика был использован из-за его высокой скорости и возможности нанесения пасты для стекла в определенных областях с определенной толщиной за один прием. Пасты, используемые с этими технологиями обработки, немного менее вязкие (Federmann et al., 2019) и содержат большую долю раствора связующего. Кроме того, раствор связующего содержит более разнообразную смесь органических добавок, чем паста, используемая для робота-дозатора, среди них: бутилгликолят, поливинилбутираль (Mowital) и неионогенное полимерное поверхностно-активное вещество (Hypermer). Условия вязкости и содержания в связующем были ранее оптимизированы для применения герметика в больших штабелях (Steinberger-Wilckens et al., 2011). Этот метод был использован для получения образцов для определения механических свойств соединений (Wei et al., 2015). Стальные подложки представляли собой стержни из Crofer22APU размером 25 × 6 × 4 мм для определения прочности на излом посредством испытаний на изгиб.
Пасты, используемые с этими технологиями обработки, немного менее вязкие (Federmann et al., 2019) и содержат большую долю раствора связующего. Кроме того, раствор связующего содержит более разнообразную смесь органических добавок, чем паста, используемая для робота-дозатора, среди них: бутилгликолят, поливинилбутираль (Mowital) и неионогенное полимерное поверхностно-активное вещество (Hypermer). Условия вязкости и содержания в связующем были ранее оптимизированы для применения герметика в больших штабелях (Steinberger-Wilckens et al., 2011). Этот метод был использован для получения образцов для определения механических свойств соединений (Wei et al., 2015). Стальные подложки представляли собой стержни из Crofer22APU размером 25 × 6 × 4 мм для определения прочности на излом посредством испытаний на изгиб.
Стальные подложки фиксируются с помощью магнитного стола, чтобы предотвратить их перемещение. Паста наносится на сетку определенной толщины и геометрии, что позволяет размещать пасту в желаемых областях при приложении давления.
После процесса образцы сушат в течение 12 ч в печи (55 ° C). Стержни были скреплены вместе с помощью приспособления для соединения, предназначенного для этой цели.
Уплотнение
Процесс герметизации был скорректирован для каждого из исследованных составов. Во всех случаях применялась статическая нагрузка для увеличения контакта между образцами, подлежащими герметизации. Стеклянную композицию 7.5B (Ba) нагревали до 1000 ° C (2 ° C / мин), стабилизацию при этой температуре в течение 10 мин, затем охлаждение до 850 ° C, стабилизацию при этой температуре в течение 10 ч и затем охлаждение до комнатная температура со скоростью 2 ° C / мин.Для стекла состава 10В (Sr) нагрев был только до 850 ° С со стабилизацией при этой температуре в течение 10 ч. В обоих случаях была промежуточная стадия при 350 ° C в течение 30 минут для выжигания всех органических компонентов, присутствующих в пастах для стекла.
Некоторые образцы были подвергнуты термоциклированию, чтобы оценить влияние этой обработки на микроструктуру герметика, чтобы оценить некоторые свойства после отжига. Программа цикла состояла из нагрева до 750 ° C (10 ° C / мин), стабилизации в течение 5 часов и охлаждения до комнатной температуры со скоростью 1 ° C / мин в течение 50 циклов.
Программа цикла состояла из нагрева до 750 ° C (10 ° C / мин), стабилизации в течение 5 часов и охлаждения до комнатной температуры со скоростью 1 ° C / мин в течение 50 циклов.
Измерения газонепроницаемости
Газонепроницаемость измерялась для образцов в форме сэндвича Crofer22APU / стеклокерамическое уплотнение / Crofer22APU и Crofer22H / стеклокерамика / Crofer22H. Эти образцы имеют углубление диаметром 10 мм в центре одной из стальных пластин, через которое газы проходят внутрь образцов. Оборудованием для обнаружения был детектор утечки гелия UL200 (Inficon), использующий перепад давления в 1 бар (da Cruz Payão Filho et al., 2000; Cela Greven, 2014).
Также было определено влияние степени кристаллизации на газонепроницаемость. Термические обработки проводили в течение 24, 100, 300 и 800 ч соответственно при 800 ° C. Газонепроницаемость соединений измерялась после термической обработки при комнатной температуре. Также оценивалось влияние термоциклирования (снова 50 циклов).
Для изучения влияния восстановительной атмосферы на это свойство и на соединения с определенной степенью кристаллизации была проведена термообработка в течение 100 часов при 800 ° C в атмосфере 4% H 2 в атмосфере Ar.
Электрическое сопротивление при высокой температуре
Электрическое сопротивление определяли с использованием сэндвич-образцов для испытаний на газонепроницаемость. Измерения проводились посредством четырехточечной проверки герметичности с использованием 4 платиновых проволок (Cela Greven et al., 2013). Сопротивление образцов при комнатной температуре превышало 2 ГОм. Измерения проводились при температуре до 850 ° C с использованием линейного нагрева 2 ° C / мин и приложения разности потенциалов 5 В. Измерения проводились в резистивной электрической печи, в которой разность потенциалов прикладывалась к образцу через трансформатор. и измерить сопротивление (Села Гревен, 2014).
Химическая совместимость уплотнений
Химическая совместимость материалов герметика со сталью и электролитом была оценена с помощью сканирующей электронной микроскопии (SEM) и химического анализа EDS поперечных сечений швов.
Для исследования микроструктуры образцов использовалось несколько электронных микроскопов. Настольная СЭМ модель TM-100 (Hitachi) использовалась для исследования поверхностей изломов и для получения изображений при малом увеличении.Модель Mira3 (Tescan) FE-SEM с сопряженным рентгеновским флуоресцентным спектрометром для дисперсной энергии (EDX) Silicon Drif Detector (SDD) -X-Max N от Oxford Instruments использовалась для наблюдений при большом увеличении. Также использовался SE Cambridge Stereoscan 360 со спектрометром EDX от Oxford Instruments.
Прочность суставов на излом
Прочность на излом герметизированных стальных стержней измерялась с использованием конфигурации четырехточечного изгиба. Соблюдалась процедура, описанная в DIN EN 843–2 (2007).Расстояние между нижней и верхней опорами составляло 40 и 20 мм соответственно. Сканирование смещения составляло 0,01 мм / мин. Испытания проводились с использованием испытательной машины Instron 1362 с датчиком нагрузки 1,5 кН (интерфейс 1210 BLR), соединенным с трубчатой печью. Смещение регистрировалось керамическим стержнем, соединенным с линейным вариационным дифференциальным трансформатором (Sangamo, LVDT, точность 1,25 мкм). Скорость нагрева составляла 2 ° C / мин, применялась предварительная нагрузка ~ 2 Н. Эти измерения проводились при комнатной и высокой температуре и на носителе из оксида алюминия.
Результаты и обсуждение
Дилатометрические свойства и вязкость стекол
О дилатометрических свойствах выбранных композиций уже сообщалось в Rodríguez-López et al. (2017). Температуры стеклования (T g ) составляют 655 ° C и 675 ° C для стекол 7,5B (Ba) и 10B (Sr) соответственно. На рисунке 1 показаны экспериментальные точки данных вязкости, скорректированные с помощью уравнений VFT и MYEGA (Mauro et al., 2009). Оба уравнения хорошо подходят.При температурах запечатывания 800–950 ° C оба состава обладают вязкостью от 10 4 до 10 7 дПа · с, поэтому можно ожидать хорошего сцепления с компонентами ячейки. Максимальная температура для герметизации ТОТЭ является спорным из-за окисления и поведение ползучести ферритных нержавеющих сталей, используемых в качестве части стека (Quadakkers и др., 2003). Испытания соединения с реактивно-воздушной пайкой проводились при температуре до 1050 ° C со временем выдержки 30 мин, что можно рассматривать как ограничивающее граничное условие для развития процесса герметизации (Weil et al., 2005).
Максимальная температура для герметизации ТОТЭ является спорным из-за окисления и поведение ползучести ферритных нержавеющих сталей, используемых в качестве части стека (Quadakkers и др., 2003). Испытания соединения с реактивно-воздушной пайкой проводились при температуре до 1050 ° C со временем выдержки 30 мин, что можно рассматривать как ограничивающее граничное условие для развития процесса герметизации (Weil et al., 2005).
Рис. 1. (A) Вязкостно-температурные кривые для стекол 7.5B (Ba) и 10B (Sr). Подбор экспериментальных точек: уравнение VFT (сплошные линии) и уравнение MYEGA (пунктирные линии). (B) Кривые вязкости в зависимости от T г / T для стекол 7,5B (Ba), 10B (Sr), оконного стекла и кремнезема, непрерывные линии показывают соответствие с уравнением MYEGA.
Стабильность уплотнения и газонепроницаемость
Робот-дозатор был использован для нанесения герметика для стекла на стальные пластины Crofer22APU и, следовательно, для получения газонепроницаемых соединений.Программа уплотнения, значения газонепроницаемости, конечная толщина уплотнения и вязкость стекла приведены в таблице 1.
Таблица 1 . Герметичность по воздуху, газонепроницаемость и конечная толщина стыков сталь / стеклокерамика / сталь.
Стеклянная композиция 10B (Sr) показала отличную газонепроницаемость 10 −9 мбар · л / с, что даже превосходит требования по газонепроницаемости для таких материалов (≤ 10 −7 мбар · л / с на см сустава; Eichler et al., 1999).Конечная толщина герметиков составляет ~ 215 мкм, что соответствует желаемому диапазону толщины штабелей Forschungszentrum Jülich (от 200 до 250 мкм; Steinberger-Wilckens et al., 2006). Напротив, композиция 7.5B (Ba) обеспечивает довольно низкую газонепроницаемость 10 -4 мбар · л / с и большую толщину ~ 430 мкм. Эта композиция обладает большей толщиной герметика из-за более высокой скорости кристаллизации и, следовательно, более высокой вязкости. Вязкости, достигнутые во время процесса герметизации, которые представлены в таблице 1, показывают хорошую смачиваемость подложки, поскольку для хорошего соединения стекло должно иметь вязкость 10 4 -10 7 дПа · с (Weil, 2006). .Вязкость стекла 7,5B (Ba) при 850 ° C (log η ~ 5,6) подходит для герметизации, но быстрая кристаллизация при этой температуре приводит к увеличению вязкости при увеличении кристаллической фракции (Meinhardt et al., 2008) и как следствие, имеется ограниченная смачиваемость подложки и большая толщина уплотнения, что приводит к плохой газонепроницаемости.
Вязкости, достигнутые во время процесса герметизации, которые представлены в таблице 1, показывают хорошую смачиваемость подложки, поскольку для хорошего соединения стекло должно иметь вязкость 10 4 -10 7 дПа · с (Weil, 2006). .Вязкость стекла 7,5B (Ba) при 850 ° C (log η ~ 5,6) подходит для герметизации, но быстрая кристаллизация при этой температуре приводит к увеличению вязкости при увеличении кристаллической фракции (Meinhardt et al., 2008) и как следствие, имеется ограниченная смачиваемость подложки и большая толщина уплотнения, что приводит к плохой газонепроницаемости.
Чтобы улучшить газонепроницаемость состава 7.5B (Ba), были использованы различные программы уплотнения, в которых также использовались более высокие температуры и нагрузки для улучшения размягчения и смачиваемости стали, способствуя кинетике уплотнения.Результаты экспериментов также приведены в таблице 1. Повышение температуры уплотнения и приложенной нагрузки значительно улучшает газонепроницаемость. Уплотнение при 950 ° C и нагрузке 1200 г улучшает скорость утечки на два порядка (10 −6 мбар · л / с), но эта кажущаяся газонепроницаемость все еще ниже требований.
Эксперименты, описанные ниже, были сосредоточены больше на повышении температуры, но в течение короткого времени соединения (10 мин), поскольку при температуре выше 1000 ° C сталь значительно размягчается.Дополнительная обработка при 850 ° C обеспечивает уплотнение с более высокой степенью кристаллизации и, таким образом, ожидается достаточная механическая стабильность при высоких температурах. Газонепроницаемость достигает 10 −7 и 10 −10 мбар · л / с при 1000 и 1050 ° C соответственно. В этих случаях значения газонепроницаемости улучшаются на три и шесть порядков соответственно, удовлетворяя требованиям газонепроницаемости (≤ 10 −7 мбар · л / с). Несмотря на отличные показатели газонепроницаемости, полученные при 1050 ° C, температура 1000 ° C была выбрана для герметизации из-за высокой диффузии хрома из стали в стеклокерамику, что следует из зеленого цвета, отображаемого стеклом. керамика, запаянная при 1050 ° C, и соответствующие образцы, которые были разбиты для проверки.
керамика, запаянная при 1050 ° C, и соответствующие образцы, которые были разбиты для проверки.
После того, как программа герметизации была выбрана, с использованием Crofer22H были подготовлены два образца типа «сэндвич» для исследования различий в газонепроницаемости и, следовательно, различий в сцеплении между ними. Такие же значения газонепроницаемости были получены при использовании разных сталей, и никаких различий в отношении сцепления, по-видимому, не существует.
Эволюция газонепроницаемости с учетом степени кристаллизации и термоциклирования
Соединения, обсуждаемые в предыдущей части, обрабатывались при температуре 800 ° C на воздухе, типичной рабочей температуре планарных ТОТЭ, и с использованием различных типов обработки, имитирующих кристаллизацию герметизирующих материалов во время работы с пакетом.Таким образом, можно реалистично изучить эволюцию газонепроницаемости с ожидаемым временем работы. Кроме того, соединения были подвергнуты термоциклированию (50 циклов), чтобы оценить влияние термоциклирования на это свойство. Во всех случаях газонепроницаемость измерялась при комнатной температуре после каждой термообработки; полученные результаты представлены в таблице 2.
Таблица 2 . Газонепроницаемость (мбар · л / с) соединений Crofer22APU / стеклокерамика / Crofer22APU в зависимости от времени обработки при 800 ° C и после термоциклирования.
Отжиг при 800 ° С в течение 800 ч не изменяет полученную газонепроницаемость состава 7.5В (Ва). Он остается постоянным в течение всего выбранного времени лечения. После термоциклирования газонепроницаемость снизилась до 1 · 10 −6 мбар · л / с, поэтому требования для этого состава не выполнялись. Уменьшение газонепроницаемости может быть результатом механических напряжений, вызванных высокими скоростями нагрева и охлаждения во время термоциклирования, что приводит к образованию небольших микротрещин.Возникновение таких трещин также может быть связано с более низким процентным содержанием стеклообразной фазы для этого состава, поскольку стеклообразная фаза потенциально способствует снятию напряжений, близких к рабочей температуре, то есть выше температуры стекла T g (Chang et al. , 2010 ; Села Гревен, 2014).
, 2010 ; Села Гревен, 2014).
Газонепроницаемость состава 10B (Sr) отличается. Этот состав соответствует требованиям газонепроницаемости для всех изученных термических обработок, хотя после 300 часов термической обработки газонепроницаемость немного снижается до примерно ≤ 5 · 10 −7 мбар · л / с, вероятно, из-за появления мелкие поры и, как следствие, низкая пористость, связанная с локальным сжатием после выделения кристаллических фаз с более высокой плотностью, чем у исходного исходного стекла.После термических циклов этот состав все еще гарантирует высокую газонепроницаемость со значениями, аналогичными тем, которые были получены после герметизации, поэтому быстрые скорости нагрева и охлаждения во время термоциклирования, по-видимому, не влияют на это свойство в этом случае. Большее количество стеклообразной фазы в этой композиции является одним из факторов, помогающих получить хорошую стабильность, поскольку она способствует релаксации напряжений. Более высокая механическая прочность этой композиции по сравнению с композицией 7.5B (Ba) (Rodríguez-López et al., 2017) является еще одним фактором, объясняющим лучшую реакцию уплотнения на термоциклирование.
Влияние атмосферы восстановителя на газонепроницаемость
В SOFC воздух используется в качестве окислителя и топлива (например, H 2 , CH 4 , биогаз, реформированное дизельное топливо) в качестве восстановителя, поэтому интересно изучить влияние высокотемпературной обработки (800 ° C). в восстановительной атмосфере на газонепроницаемость стыков. В этом случае использовалась однократная термообработка в течение 100 ч в атмосфере Ar с 4% H 2 . Отобранные образцы представляли собой соединения, которые изначально были герметизированы на воздухе (в соответствии с оптимизированными процедурами герметизации, описанными в экспериментальной части) с определенной степенью кристаллизации (24 и 100 ч при 800 ° C).Введение восстановительной атмосферы в ячейку обычно проводят после того, как уплотнение приобрело определенную кристаллизацию (после завершения программы герметизации), которая, как ожидается, обеспечит подходящую механическую стабильность.
Состав 10B (Sr) обеспечивает отличную газонепроницаемость, полученную для всех образцов, обработанных в восстановительной атмосфере (≤ 2 · 10 −9 мбар · л / с), что подтверждает высокую стабильность в этой среде. Напротив, композиция 7.5B (Ba) показала снижение значений газонепроницаемости, что привело к значениям (10 -5 -10 -6 ), меньшим, чем требуемые, и, как следствие, эта композиция обладает низкой стабильностью. в атмосфере водорода.
Кристаллические фазы после процесса герметизации
Высокотемпературная дифракция рентгеновских лучей использовалась во время термической обработки, имитирующей программу герметизации.
На рис. 2А показаны результаты для состава стекла 7,5В (Ва). Первое сканирование проводится через 1 мин после достижения 1000 ° C; температура не стабилизировалась в течение длительного времени, так как используется короткая герметизирующая обработка (10 мин). На дифрактограмме при 1000 ° C показаны первые кристаллические фазы BaSi 2 O 5 , BaMg 2 Si 2 O 7 и Ba 3 Si 5 O 13 .Результаты показывают, что BaSi 2 O 5 является основной фазой при всех исследованных температурах и временах, хотя пики фазы BaMg 2 Si 2 O 7 также увеличиваются с точки зрения ее интенсивности, являясь второй преобладающей. фаза. При увеличении времени кристаллизации фаза Ba 3 Si 5 O 13 исчезает и появляется как новая фаза Ba 5 Si 8 O 21 . После завершения термообработки фазы: BaSi 2 O 5 , BaMg 2 Si 2 O 7 и Ba 5 Si 8 O 21 .Это те же самые фазы, определенные Родригес-Лопесом и др. (2017) для постоянной обработки при 800 ° C (без нагрева до 1000 ° C).
Рис. 2. (A) Высокотемпературная дифракция рентгеновских лучей состава 7.5B (Ba). Скорость нагрева 2 ° C / мин и скорость сканирования 2θ: 7 ° / мин при 1000 ° C и 800 ° C — 0 ч и 2 ° / мин для остальных сканирований. (B) Высокотемпературная XRD состава 10B (Sr). Скорость нагрева: 2 ° C / мин и скорость сканирования 2θ: 0,5 ° / мин.
2. (A) Высокотемпературная дифракция рентгеновских лучей состава 7.5B (Ba). Скорость нагрева 2 ° C / мин и скорость сканирования 2θ: 7 ° / мин при 1000 ° C и 800 ° C — 0 ч и 2 ° / мин для остальных сканирований. (B) Высокотемпературная XRD состава 10B (Sr). Скорость нагрева: 2 ° C / мин и скорость сканирования 2θ: 0,5 ° / мин.
На рис. 2B показаны результаты XRD при высокой температуре для состава стекла 10B (Sr) с использованием программы герметизации, описанной в экспериментальной части.
После достижения 850 ° C могут наблюдаться пики, соответствующие фазе Sr 2 MgSi 2 O 7 , и есть другие дифракционные пики, отнесение которых невозможно, как уже объяснялось в Rodríguez-López et al. (2017). Интенсивность пиков увеличивается со временем обработки, являющейся преобладающей фазой Sr 2 MgSi 2 O 7 после завершения обработки герметизацией. После термической обработки дифракционные пики совпадают с пиками, полученными в работе Rodríguez-López et al.(2017), поэтому нет разницы в выделенной кристаллической фазе при использовании более высокой температуры термообработки.
Электрическое сопротивление соединений при высокой температуре
Электрическое сопротивление после герметизации при высокой температуре оценивали с использованием четырехточечной схемы. На рис. 3А показано сопротивление как функция температуры для двух охарактеризованных составов стекла. Увеличение, показанное на вставке, показывает, что обе композиции обеспечивают хорошие изолирующие свойства, хотя стронциевый состав в целом приводит к более высокому сопротивлению при температурах испытаний.
Рис. 3. (A) Электрическое сопротивление соединений композиций 7,5B (Ba) и 10B (Sr) после герметизации и при высокой температуре. (B) Электрическое сопротивление при высокой температуре соединений состава 10B (Sr) после герметизации и после 300 ч термообработки при 800 ° C.
Все соединения показали электрическое сопротивление при комнатной температуре выше 2 ГОм, сопротивление уменьшается с повышением температуры.
В таблице 3 представлено удельное сопротивление каждого соединения при различных температурах.Требования к этим материалам — удельное сопротивление ≥10 3 Ом · м при типичной рабочей температуре батареи 800 ° C, поэтому оба состава удовлетворяют требованиям для этих материалов. Композиция 10B (Sr) имеет более высокое сопротивление при температурах, представленных в таблице 3, хотя значения удельного сопротивления имеют одинаковый порядок величины для обоих составов, за исключением температуры 700 ° C, где композиция 10B (Sr) обладает изоляционным характером.
Таблица 3 . Электрическое сопротивление при высокой температуре стыков.
Состав с лучшими изолирующими свойствами был выбран для оценки эволюции этого свойства со степенью кристаллизации. Для этого был выбран стык со стеклом состава 10B (Sr) с выдержкой 300 ч при 800 ° C. На рис. 3В представлена разница удельных сопротивлений между соединением после герметизации и другим соединением после 300 часов термического старения. В случае состаренного образца наблюдается сильное увеличение удельного сопротивления. В таблице 3 также показаны значения удельного сопротивления обоих образцов при различных температурах испытаний.Старый образец показывает значения 185 МОм · м при 800 ° C, в то время как соединение сразу после герметизации имеет удельное сопротивление 38 МОм · м при той же температуре. Таким образом, увеличение удельного сопротивления примерно на один порядок подтверждается при увеличении степени кристаллизации.
Кристаллические фазы не только обеспечивают механическую стабильность при высокой температуре, но также обеспечивают более высокие изолирующие свойства, следовательно, улучшая электрические характеристики пакета.
Химическая совместимость суставов
Ранее описанные соединения были заделаны, вырезаны и отполированы для исследования микроструктуры с помощью SEM и EDS. Для изучения химической совместимости были выбраны соединения Crofer22APU-стеклокерамика-Crofer22APU после герметизации и после 800 ч термообработки при 800 ° C на воздухе. Таким образом, можно было наблюдать за развитием соединений и микроструктуры стеклокерамики от процесса герметизации до короткого периода обработки, имитирующего работу пакета. Были также выбраны образцы, подвергнутые термическому циклу, и соединения, обработанные в пониженных условиях (4% H 2 / Ar), чтобы изучить, как эти два явления влияют на соединения и микроструктуры.
Состав 7.5B (Ba)
На рис. 4 показано поперечное сечение шва с составом 7.5B (Ba) после программы герметизации. Пустота, наблюдаемая на изображениях, является результатом разрушения образца для проверки после герметизации. На рис. 4А представлено общее изображение, на котором можно наблюдать внешнюю и центральную зоны стыка. Наблюдается высокая пористость во внешних зонах, вероятно, из-за недостаточного испарения связующего, используемого для нанесения стеклянного порошка.Стеклокерамика демонстрирует хорошее уплотнение и высокую степень кристаллизации (Рисунки 4B, C) после герметизации, обнаруженные небольшие трещины могут быть причиной того, что этот состав не достиг высокой газонепроницаемости по сравнению с составом 10B (Sr), хотя они также может быть связано с механическим раскрытием образца, например, с этим процессом может быть связана большая поперечная трещина (рис. 4В). Детальное изображение границы раздела между стеклокерамикой и сталью Crofer22APU демонстрирует отличное сцепление с наличием кристаллизации на границе раздела.
Рисунок 4 . СЭМ-изображения стыка Crofer22APU-стеклокерамика 7.5B (Ba) после герметизации (черная пустота из-за открытия образца для проверки). (A) Общее изображение сустава, показывающее внешнюю и центральную области. (B) Деталь стыка стали и герметика. (C) Микроструктура уплотнения и элементный анализ EDX.
Точечный анализ EDX (рис. 4C, таблица 4) показывает структуру и состав, очень похожие на те, что представлены у Rodríguez-López et al.(2017) для стеклокерамики после 24 ч термообработки при 800 ° C. Спектры 1 и 2 указывают на присутствие Ba, Si и O, соответствующих силикату бария BaSi 2 O 5 , хотя идентификация с помощью EDX различных присутствующих силикатов бария в этой стеклокерамике затруднена из-за их близкой стехиометрии. . Спектры 3 и 4 соответствуют фазе с удлиненными кристаллами и более темным контрастом, содержащей Mg: BaMg 2 Si 2 O 7 .Наконец, спектры 5 и 6 соответствуют оставшейся стекловидной фазе.
Таблица 4 . Элементный анализ, соответствующий рисунку 4.
Граница раздела сталь / стеклокерамика была исследована более подробно с помощью линейного сканирования EDX (не показано). Элементный анализ не выявляет наличия какого-либо промежуточного слоя между стеклокерамикой и сталью, хотя имеется небольшая диффузия стальных элементов в сторону стеклокерамики, что указывает на то, что образованный самородным слоем оксида хрома растворяется во время процесса герметизации, обеспечивая подходящее соединение.
Состав 10B (Sr)
Фиг. 5A представляет собой общий вид поперечного сечения соединения композиции 10B (Sr) после программы герметизации, где можно заметить, что образец обладает закрытой пористостью и более низкой степенью кристаллизации, чем композиция 7.5B (Ba). . Если мы сравним микроструктуру стеклокерамики в стыке с микроструктурой стеклокерамики после 24 часов термообработки при 750 ° C, повышение на 100 ° C приводит к явному увеличению кристаллизации, а также размер кристаллитов (Rodríguez-López et al., 2017). На рисунках 5B, C показаны детали стыка уплотнение-сталь и очень хорошее сцепление между стеклокерамикой и Crofer22APU с ростом кристаллов, начиная с границы раздела. Поверхность стали, кажется, усиливает зародышеобразование. На рисунках 5C, D и в таблице 5 показаны зоны точечного анализа с помощью EDX. Спектры 1 и 2 соответствуют фазе Sr 2 MgSi 2 O 7 с аналогичным соотношением Si и Sr. Спектры 3 и 4 соответствуют фазе со стехиометрией SrMgSi 2 O 6 , образованной кристаллы с темным контрастом и большего размера, чем на предыдущем этапе.Темные точки небольшого размера соответствуют спектрам 5 и 6, которые представляют обогащение SiO 2 . Спектры 7 и 8 соответствуют оставшейся стеклообразной фазе, а 9 — стали Crofer22APU. Спектры 10 и 11 соответствуют кристаллам фазы Sr 2 MgSi 2 O 7 вблизи границы раздела стали. Кристаллиты в этой области ближе к стали показывают небольшое присутствие Cr и Fe, которые диффундировали из стали в стеклокерамику.
На рисунках 5C, D и в таблице 5 показаны зоны точечного анализа с помощью EDX. Спектры 1 и 2 соответствуют фазе Sr 2 MgSi 2 O 7 с аналогичным соотношением Si и Sr. Спектры 3 и 4 соответствуют фазе со стехиометрией SrMgSi 2 O 6 , образованной кристаллы с темным контрастом и большего размера, чем на предыдущем этапе.Темные точки небольшого размера соответствуют спектрам 5 и 6, которые представляют обогащение SiO 2 . Спектры 7 и 8 соответствуют оставшейся стеклообразной фазе, а 9 — стали Crofer22APU. Спектры 10 и 11 соответствуют кристаллам фазы Sr 2 MgSi 2 O 7 вблизи границы раздела стали. Кристаллиты в этой области ближе к стали показывают небольшое присутствие Cr и Fe, которые диффундировали из стали в стеклокерамику.
Рисунок 5 .СЭМ-изображения стыка Crofer22APU-стеклокерамическое уплотнение 10B (Sr) после герметизации при 850 ° C в течение 10 часов. (A) Общее изображение шва, показывающее внешнюю и центральную области. (B, C) Деталь стыка стали и герметика. (C, D) Микроструктура пломбы и точечный элементный анализ методом EDX.
Таблица 5 . Элементный анализ, соответствующий рисунку 5.
Линейный элементный анализ, полученный для обеих границ раздела между сталью и уплотнением после соединения (не показан), показывает, что не происходит образования нового слоя и диффузии Fe и Cr из стали.В случае нижней границы раздела также наблюдается хорошее сцепление, но наблюдается значительная диффузия Cr из стали до ~ 3 мкм в уплотнение.
Элементный анализ EDX на границах раздела сталь-стеклокерамика, обработанный при 800 ° C в течение 800 ч (рисунки 6A – D), показывает в обоих случаях наличие слоя, обогащенного Cr, Mn и O, толщиной ~ 2–3 мкм. толщина. Образование этого слоя указывает на возможное образование (Mn, Cr) 3 O 4 , образование этого слоя широко описано в литературе (Mantel, 2000; Yang et al. , 2003, 2004; Чоу и др., 2008; Smeacetto et al., 2009, 2013; Lin et al., 2012; Kaur et al., 2013) на поверхности ферритных нержавеющих сталей, состоящих из Fe, Cr и Mn, таких как Crofer22APU. Этот слой является стабильным покрытием (Kaur et al., 2013) и менее реактивным, чем Cr 2 O 3 , который также может образовываться на поверхности стали. Наличие этого слоя предотвращает испарение хрома и, следовательно, сводит к минимуму коррозию стали, а также предотвращает возможное образование хроматов стронция с высоким коэффициентом теплового расширения TEC (21–23 · 10 −6 K −1 , 25– 1000 ° C; Chou et al., 2007), что может вызвать физическое разделение уплотнения и стали. Другой положительный аспект этого слоя заключается в том, что он улучшает герметизирующую поверхность, действуя как переходный слой между металлом и стеклокерамическим уплотнением.
, 2003, 2004; Чоу и др., 2008; Smeacetto et al., 2009, 2013; Lin et al., 2012; Kaur et al., 2013) на поверхности ферритных нержавеющих сталей, состоящих из Fe, Cr и Mn, таких как Crofer22APU. Этот слой является стабильным покрытием (Kaur et al., 2013) и менее реактивным, чем Cr 2 O 3 , который также может образовываться на поверхности стали. Наличие этого слоя предотвращает испарение хрома и, следовательно, сводит к минимуму коррозию стали, а также предотвращает возможное образование хроматов стронция с высоким коэффициентом теплового расширения TEC (21–23 · 10 −6 K −1 , 25– 1000 ° C; Chou et al., 2007), что может вызвать физическое разделение уплотнения и стали. Другой положительный аспект этого слоя заключается в том, что он улучшает герметизирующую поверхность, действуя как переходный слой между металлом и стеклокерамическим уплотнением.
Рис. 6. (A) Увеличение границы раздела улучшенная сталь Crofer22APU / стеклокерамика и линейный анализ (B) EDX. (C) Увеличение границы раздела низшая сталь / стеклокерамика и линейный анализ (D) EDX.Стеклокерамика состава 10В (Sr), выдержанная при 800 ° С в течение 800 ч.
Картирование границы раздела нижняя сталь-стеклокерамика (рис. 7) подтверждает присутствие ранее описанного слоя шпинели, поскольку Cr и Mn сосредоточены между сталью и стеклокерамикой. Нет диффузии стальных элементов через слой, что подтверждает устойчивость этого слоя к разрушению стали. Картирование также позволяет идентифицировать некоторые кристаллические фазы, такие как фаза со стехиометрией SrMgSi 2 O 6 с более высоким содержанием Mg, которое соответствует зонам высокой концентрации этого элемента.Зоны с высоким содержанием Si соответствуют фазе достижения SiO 2 . Хотя Sr распределен во всех проанализированных областях, можно выделить зоны с более высокой концентрацией этого элемента, соответствующие фазе Sr 2 MgSi 2 O 7 .
Рисунок 7 . Картирование элементов EDX для нижнего стыка сталь-стеклокерамика 10B (Sr), обработанного при 800 ° C в течение 800 часов.
Поперечное сечение образцов, обработанных в восстановительной атмосфере и после термоциклирования, показало аналогичные хорошие характеристики сцепления (рисунки не показаны).Образцы, подвергнутые термоциклированию, как и ожидалось, показали более высокую степень кристаллизации, но стабильное соединение.
Прочность суставов на излом
Для определения напряжения разрушения стыков при комнатной и высокой температуре стальные стержни Crofer22APU герметизировали стеклами двух выбранных составов в соответствии с экспериментальной процедурой и с оптимизированными программами герметизации.
После герметизации образцы были ректифицированы для получения двух плоских поверхностей и подверглись механическим испытаниям с использованием 4-точечного изгиба.Затем определяли напряжение разрушения для образцов с различной толщиной уплотнения.
На рис. 8 показаны кривые «нагрузка-смещение» в зависимости от температуры и толщины герметика. Стержни, запечатанные и испытанные при комнатной температуре, имеют линейную кривую нагружения вплоть до разрушения, что демонстрирует хрупкое поведение этого типа материалов; такое поведение наблюдается для одного слоя, нанесенного методом трафаретной печати, а также для двух слоев стеклокерамики. Линейное поведение исчезает при 800 ° C, и образцы разрушаются при более низких нагрузках по сравнению с результатами измерений при комнатной температуре.
Рисунок 8 . Кривые нагрузки-деформации для герметизации стержней с одним или двумя слоями, нанесенными методом трафаретной печати, в зависимости от температуры испытания для композиций 7,5B (Ba) и 10B (Sr).
Фактически, состав 10B (Sr), характеризуемый при 700 ° C, показывает кривую с нелинейным трендом, где пластическая деформация продолжается в течение длительного времени нагружения, таким образом, достигаются высокие деформации, и соединение в конце концов разрушается при более высокие значения нагрузки. В этом случае вязкоупруго-пластическое течение, находящееся под влиянием оставшейся стеклообразной фазы, способствует пластичному поведению при температурах выше T g (Chang et al., 2010; Zhao, 2013), действующий как механизм усиления сустава и указывающий также на эффект релаксации напряжений из-за присутствия стеклообразной фазы (Chang et al., 2010). Такое поведение происходит независимо от толщины герметика и указывает на возможный эффект «самозаживления» трещин, образовавшихся во время испытания из-за размягчения стеклообразной фазы, что объясняет более высокое механическое сопротивление этих образцов.
В этом случае вязкоупруго-пластическое течение, находящееся под влиянием оставшейся стеклообразной фазы, способствует пластичному поведению при температурах выше T g (Chang et al., 2010; Zhao, 2013), действующий как механизм усиления сустава и указывающий также на эффект релаксации напряжений из-за присутствия стеклообразной фазы (Chang et al., 2010). Такое поведение происходит независимо от толщины герметика и указывает на возможный эффект «самозаживления» трещин, образовавшихся во время испытания из-за размягчения стеклообразной фазы, что объясняет более высокое механическое сопротивление этих образцов.
Для оценки происхождения массивных деформаций образцов состава 10B (Sr) при высокотемпературных измерениях из-за их более низкой тугоплавкости по сравнению с составом 7.5B (Ba) вязкость измерялась с использованием «изгиба пучка» стеклокерамических брусков с той же термической обработкой (850 ° C в течение 10 ч; график не показан; Rodríguez-López, 2016). Наблюдалось сильное увеличение вязкости стеклокерамики, поэтому от массивного размягчения этого материала отказываются как при 700 ° C, так и при 800 ° C, поскольку он обладает высокой вязкостью от 10 12 до 10 9,5 дПа · с в диапазон температур от 938 до 950 ° C.
В таблице 6 приведены значения прочности на изгиб, полученные в зависимости от толщины уплотнения и температуры испытания.Состав 7.5B (Ba) обеспечивает среднее сопротивление 42 МПа при комнатной температуре. Эти значения превосходят полученные для состава 10B (Sr) со значениями 30 МПа. Вероятно, это связано с более высокой температурой герметизации бариевой композиции, которая обеспечивает лучшее сцепление между сталью и стеклокерамикой, хотя это также приводит к более высокому разрушению стали.
Таблица 6 . Сопротивление изгибу при комнатной и высокой температуре в зависимости от толщины уплотнения.
Оба уплотнения демонстрируют снижение сопротивления изгибу с повышением температуры со снижением значений на ~ 75% для состава бария и на ~ 66% для состава стронция при 800 ° C, обеспечивая значения 11 и 10 МПа соответственно. . Более высокие значения получены для стронциевого состава при 700 ° C с сопротивлением 65 МПа из-за уже упомянутого усиливающего эффекта текучести стеклообразной фазы. Такое поведение свидетельствует о положительном влиянии наличия стеклообразной фазы на механические свойства в данном случае.
. Более высокие значения получены для стронциевого состава при 700 ° C с сопротивлением 65 МПа из-за уже упомянутого усиливающего эффекта текучести стеклообразной фазы. Такое поведение свидетельствует о положительном влиянии наличия стеклообразной фазы на механические свойства в данном случае.
Влияние толщины герметика показывает слегка отрицательную тенденцию к этому механическому свойству в случае состава 7.5B (Ba), хотя значения постоянны в пределах погрешности измерения. Однако для состава 10B (Sr) это влияние заметно отрицательно, уменьшая значения прочности при большей толщине. Это влияние можно объяснить сжимающими напряжениями, возникающими при охлаждении, которые вызваны различиями в TEC между стеклокерамикой и сталью, которые вызывают увеличение тягового сопротивления.Остаточное напряжение сжатия тем выше, чем выше разница ПЭС (Malzbender et al., 2014). Этот эффект более важен для состава стронция, как видно на рисунке 9A, из-за большей разницы в TEC между стеклокерамикой и Crofer22APU (Rodríguez-López et al., 2017). Согласно этому аргументу, стеклокерамика 7.5B (Ba) должна иметь меньшую разницу в TEC с Crofer22APU, что приведет к более низким остаточным напряжениям в соединении.
Рисунок 9.(A) Влияние толщины уплотнения на значения сопротивления изгибу, левый состав 7.5B (Ba), правый состав 10B (Sr). (B) Распределение Вейбулла для стержней, соединенных одним или двумя слоями стеклокерамики 7.5B (Ba).
Распределение Вейбулла использовалось для проверки достоверности этих значений из-за большего количества протестированных образцов состава бария и с учетом релевантности количества тестов при использовании этого распределения только результаты этого состава использовались для статистического анализа. анализ.На рис. 9В показаны результаты двухпараметрического распределения Вейбулла для стержней, запечатанных одним и двумя слоями стеклокерамики 7,5В (Ва). Обратите внимание: хотя здесь и используется для обсуждения, строго статистический анализ требует большего количества образцов ~ 30, поэтому представленный здесь анализ используется только для выявления тенденций в поведении материалов.
Модули Вейбулла находятся в диапазоне от 5,7 до 6,1, с небольшим увеличением надежности образцов большей толщины, однако, учитывая ограниченное количество образцов и статистическую погрешность, они могут считаться идентичными; эти значения аналогичны значениям, полученным для спеченных прутков этого состава после 24 часов обработки (Rodríguez-López et al., 2017), который показал значение 6,4.
Как и ожидалось, значения прочности соединений ниже, чем у насыпных материалов из-за различий в условиях спекания и обработки, использования связующего, пластификатора и т. Д., Которые влияют на конечную микроструктуру стеклокерамики и, в частности, на ее пористость.
Если полученные значения сравнить с теми, которые описаны в литературе с использованием того же типа испытания с аналогичной толщиной герметика, значения 42 МПа, полученные при комнатной температуре для состава 7.5B (Ba) превосходят описанные в литературе значения 34 и 25 МПа для уплотнений HP и B (Zhao, 2013; Malzbender et al., 2014), соответственно, и аналогичны значениям 30 МПа, полученным для состав 10Б (Sr). При 800 ° C значение ~ 10 МПа, полученное для обоих составов, в 10 раз выше, чем полученное для уплотнения H-P со значениями ~ 1 МПа, но меньше, чем значение, представленное уплотнением B со значениями ~ 30 МПа. Тем не менее, лучшие значения сопротивления изгибу (65 МПа), полученные при 700 ° C для состава 10B (Sr), превосходят таковые у уплотнений, испытанных в аналогичной конфигурации.
Выводы
Путем оптимизации программ герметизации были получены газонепроницаемые соединения двух стеклокерамических композиций, которые удовлетворяют требованиям для работы ТОТЭ. Композиция 7,5B (Ba) требует более высокой температуры уплотнения (1000 ° C) для достижения соответствующей газонепроницаемости из-за более высокой скорости кристаллизации, высокой вязкости и плохой смачиваемости. Температура герметизации этой композиции является недостатком для ее использования в качестве герметизирующего материала IT-SOFC, поскольку сталь может подвергаться большему разложению и даже может размягчаться при этих температурах. Напротив, композиция 10B (Sr) обладает достаточной газонепроницаемостью, когда она герметизируется при температуре, типичной для этих материалов (850 ° C). Для этого состава значения газонепроницаемости соответствуют требованиям во всех изученных случаях: изменение во времени термической обработки на воздухе (степень кристаллизации уплотнения), реакция термоциклирования и термическая обработка в восстановительной атмосфере. Бариевый состав имеет низкую стабильность газонепроницаемости, показывая значения ниже требуемых после термоциклирования и термической обработки в восстановительной атмосфере.
Напротив, композиция 10B (Sr) обладает достаточной газонепроницаемостью, когда она герметизируется при температуре, типичной для этих материалов (850 ° C). Для этого состава значения газонепроницаемости соответствуют требованиям во всех изученных случаях: изменение во времени термической обработки на воздухе (степень кристаллизации уплотнения), реакция термоциклирования и термическая обработка в восстановительной атмосфере. Бариевый состав имеет низкую стабильность газонепроницаемости, показывая значения ниже требуемых после термоциклирования и термической обработки в восстановительной атмосфере.
Кристаллические фазы, полученные во время и после запечатывания, не изменяются при использовании более высоких температур, чем те, которые используются для получения объемной стеклокерамики из стеклянного порошка. Заключительные фазы: BaSi 2 O 5 , BaMg 2 Si 2 O 7 , Ba 3 S 5 O 13 и Ba 5 Si 8 O 21 для состава 7.5B (Ba) и Sr 2 MgSi 2 O 7 для состава 10B (Sr).Основные кристаллические фазы не были изменены, поэтому использование различных температур не стимулирует осаждение новых кристаллических фаз.
Высокотемпературное электрическое сопротивление соединений соответствует электрическим требованиям, предъявляемым к уплотнениям ТОТЭ. Состав 10B (Sr) показал более высокое удельное сопротивление, чем состав бария. Кристаллические фазы способствуют изолирующему характеру, поскольку увеличение электрического сопротивления происходит после термического старения.
Исследование поперечного сечения запечатанных образцов показало большую стабильность соединений 10B (Sr) из-за образования шпинели Cr и Mn, которая придает стабильность границе раздела.Этот слой уменьшает испарение Cr из стали и диффузию стальных элементов в стеклокерамическое уплотнение, а также помогает минимизировать образование продуктов коррозии. Хотя образование этого слоя не наблюдается после герметизации, его обнаруживают после термообработки в восстановительной атмосфере, на воздухе и после термоциклирования. Этот слой не появляется в случае соединений 7.5B (Ba), где обнаружена диффузия Cr из стали в стеклокерамику.
Хотя образование этого слоя не наблюдается после герметизации, его обнаруживают после термообработки в восстановительной атмосфере, на воздухе и после термоциклирования. Этот слой не появляется в случае соединений 7.5B (Ba), где обнаружена диффузия Cr из стали в стеклокерамику.
Состав 10B (Sr) обеспечивает большую стабильность соединений при термоциклировании из-за его более высокого содержания в остаточной стеклообразной фазе, которая помогает снять напряжения, возникающие во время термоциклирования.
Значения прочности на изгиб, полученные при комнатной температуре (42 МПа), были выше для композиции 7.5B (Ba) из-за превосходного уплотнения этой композиции в центре швов. Значения прочности при 800 ° C показывают снижение на 66–75% (~ 10 МПа) по сравнению с результатами при комнатной температуре для обоих составов. Композиция 10B (Sr) демонстрирует лучшие результаты по прочности при 700 ° C со значением 65 МПа. Кривые нагрузка-смещение демонстрируют линейное поведение при комнатной температуре, которое трансформируется в нелинейное поведение при повышении температуры испытания.При 700 ° C кривые состава 10B (Sr) демонстрируют пластичность, что указывает на эффект вязкой деформации остаточной стеклообразной фазы. Это явление увеличивает прочность соединения, вероятно, из-за самозаживления трещин, вызванных во время испытаний вязкоупругой / пластической деформацией стеклообразной фазы. Значение, полученное при 700 ° C, выше, чем значения, найденные в литературе для уплотнительных материалов ТОТЭ.
В целом, результаты подтверждают, что разработанный и испытанный материал является многообещающим для долгосрочного стабильного применения SOFC и SOEC в усовершенствованных конструкциях батарей, способствующих увеличению срока службы в термоциклических условиях.
Заявление о доступности данных Необработанные данные, подтверждающие выводы этой статьи, будут предоставлены авторами без излишних оговорок любому квалифицированному исследователю.
Авторские взносы
SR-L выполнил большую часть экспериментальных работ и участвовал в написании и обсуждении статьи. JM провел измерения прочности суставов и участвовал в обсуждении результатов и написании статьи. VJ и FS провели эксперименты по высокотемпературной дифракции рентгеновских лучей и участвовали в обсуждении результатов.Компания SG-B координировала эксперименты по газонепроницаемости и электрическому сопротивлению и участвовала в обсуждении результатов. MP разработал и структурировал работу, координируя все эксперименты, и участвовал в написании и обсуждении статьи.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы благодарят европейский проект FP7-JTI-CP-FCH, Работа в направлении массового производства, недорогих и надежных стеков SOFC (MMLRC = SOFC), номер проекта: 278525.Авторы также благодарны проектам CNPq / PVE 400590/2013 и CAPES за финансовую поддержку и C-LABMU / UEPG за использование исследовательской базы. SR-L благодарит финансовую поддержку, предоставленную Trust of the Journal of the European Ceramic Society (JECS Trust) в рамках гранта № 201354.
Список литературы
Блюм, Л., Гросс, С. М., Мальцбендер, Дж., Пабст, У., Пексен, М., Петерс, Р. и др. (2011). Исследование герметичности твердооксидных топливных элементов в условиях, связанных с дымовой трубой, в Forschungszentrum Jülich. Дж . Источники энергии 196, 7175–7181. DOI: 10.1016 / j.jpowsour.2010.09.041
CrossRef Полный текст | Google Scholar
Села Гревен, Б. (2014). Армирующий стеклокерамический герметик для высокотемпературных применений (кандидатская диссертация). Forschungszentrum Jülich, RWTH Ахенский университет.
Google Scholar
Села Гревен, Б., Гросс-Барсник, С. М., Федерманн, Д., Конрад, Р. (2013). Оценка прочности многослойных стеклокерамических герметиков. Топливные элементы 13, 565–571. DOI: 10.1002 / fuce.201200155
М., Федерманн, Д., Конрад, Р. (2013). Оценка прочности многослойных стеклокерамических герметиков. Топливные элементы 13, 565–571. DOI: 10.1002 / fuce.201200155
CrossRef Полный текст | Google Scholar
Чанг, Х. Т., Линь, К. К. и Лю, К. К. (2010). Влияние кристаллизации на высокотемпературные механические свойства стеклянного герметика для твердооксидных топливных элементов. J. Источники энергии 195, 3159–3165. DOI: 10.1016 / j.jpowsour.2009.12.008
CrossRef Полный текст | Google Scholar
Чоу, Ю.С., Стивенсон, Дж. У., и Сингх, П. (2007).Новые тугоплавкие уплотнительные стекла из силиката щелочноземельных металлов для плоских твердооксидных топливных элементов. J. Electrochem. Soc . 154, B644 – B651. DOI: 10,1149 / 1,2733868
CrossRef Полный текст | Google Scholar
Чоу, Ю.С., Стивенсон, Дж. У., и Сингх, П. (2008). Влияние предварительного окисления и старения в окружающей среде на прочность уплотнения нового герметизирующего стекла для высокотемпературных твердооксидных топливных элементов (ТОТЭ) с металлическим соединением. J. Источники энергии 184, 238–244. DOI: 10.1016 / j.jpowsour.2008.06.020
CrossRef Полный текст | Google Scholar
да Круз Паяо Филью, Дж., Шмидт, В., и Шредер, Г. (2000). Fundamentos de Ensaio de Vazamento e Estanqueidade . Юлих: Forschungszentrum Jülich.
Google Scholar
DIN EN 843–2, 2007–03. (2007). Передовая техническая керамика — Механические свойства монолитной керамики при комнатной температуре — Часть 2: Определение модуля Юнга, модуля сдвига и коэффициента Пуассона , (Берлин).
Google Scholar
Эйхлер, К., Солоу, Г., Отчик, П., и Шаффрат, В. (1999). BAS (BaO-Al 2 O 3 -SiO 2 ) — стекла для высокотемпературного применения. J. Eur. Ceram. Soc . 19, 1101–1104. DOI: 10.1016 / S0955-2219 (98) 00382-3
CrossRef Полный текст | Google Scholar
Федерманн Д. , Зерфасс Х. и Гросс-Барсник С. (2019). Метод изготовления зеленой печати из припоя. Патент США № 10,170,775 B2 .Вашингтон, округ Колумбия: Патент США и Ведомство по товарным знакам.
, Зерфасс Х. и Гросс-Барсник С. (2019). Метод изготовления зеленой печати из припоя. Патент США № 10,170,775 B2 .Вашингтон, округ Колумбия: Патент США и Ведомство по товарным знакам.
Google Scholar
Фергус, Дж. У. (2005). Герметики для твердооксидных топливных элементов. Дж . Источники энергии 147, 46–57. DOI: 10.1016 / j.jpowsour.2005.05.002
CrossRef Полный текст | Google Scholar
Haanappel, V.A.C., Shemet, V., Gross, S.M., Koppitz, T., Menzler, N.H., Zahid, M., et al. (2005). Поведение различных стеклокерамических герметиков с ферритными сталями в условиях моделирования батареи ТОТЭ. J Источники энергии 150, 86–100. DOI: 10.1016 / j.jpowsour.2005.02.015
CrossRef Полный текст | Google Scholar
ISO 7884–2. (1987). Glass — Вязкость и фиксированные точки вязкости — Часть 2: Определение вязкости ротационными вискозиметрами , (Женева).
Google Scholar
Каур Г., Сингх К., Пандей О. П., Хома Д., Скотт Б. и Пикрелл Г. (2013). Структурные и термические свойства стеклокомпозитных уплотнений и их химическая совместимость с Crofer 22APU для применения в твердооксидных топливных элементах. J. Источники энергии 240, 458–470. DOI: 10.1016 / j.jpowsour.2013.03.142
CrossRef Полный текст | Google Scholar
Лессинг П. А. (2007). Обзор технологий герметизации, применимых к твердооксидным электролизерам. Дж . Mater. Sci . 42, 3465–3476. DOI: 10.1007 / s10853-006-0409-9
CrossRef Полный текст | Google Scholar
Лин, К. К., Чен, Дж. Й., Тиан, Дж. У., Чанг, Л. К., и Ву, С. Х. (2012). Прочность соединения стеклокерамического герметика для твердооксидных топливных элементов с металлическими межсоединениями. J. Источники энергии 2015, 307–317. DOI: 10.1016 / j.jpowsour.2012.01.048
CrossRef Полный текст | Google Scholar
Лин, К. К., Лю, Ю. А., Ву, С. Х., Лю, К. К., и Ли, Р. Ю. (2015). Прочность соединения стеклокерамического герметика для твердооксидных топливных элементов с металлическими межсоединениями в восстановительной среде. J. Источники энергии 280, 272–288. DOI: 10.1016 / j.jpowsour.2015.01.126
Прочность соединения стеклокерамического герметика для твердооксидных топливных элементов с металлическими межсоединениями в восстановительной среде. J. Источники энергии 280, 272–288. DOI: 10.1016 / j.jpowsour.2015.01.126
CrossRef Полный текст | Google Scholar
Мальцбендер, Дж., Чжао, Ю.и Бек Т. (2014). Разрушение и ползучесть стеклокерамических твердооксидных герметиков для топливных элементов. J. Источники энергии 246, 574–580. DOI: 10.1016 / j.jpowsour.2013.08.010
CrossRef Полный текст | Google Scholar
Мантел, М. (2000). Влияние двойного оксидного слоя на герметизацию металл – стекло. J. Non. Cryst. Твердые тела 273, 294–301. DOI: 10.1016 / S0022-3093 (00) 00137-X
CrossRef Полный текст | Google Scholar
Мауро, Дж. К., Юэ, Ю., Эллисон, А.Дж., Гупта П. К. и Аллан Д. К. (2009). Вязкость стеклообразующих жидкостей. Natl. Акад. Sci. США А . 106: 19780. DOI: 10.1073 / pnas.0
5106PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мейнхардт, К. Д., Ким, Д. С., Чоу, Ю. С., и Вейл, К. С. (2008). Синтез и свойства стеклокерамического герметика для твердооксидных топливных элементов из алюмосиликата бария. J. Источники энергии 182, 188–196. DOI: 10.1016 / j.jpowsour.2008.03.079
CrossRef Полный текст | Google Scholar
Нильсен, К.А., Солванг, М., Нильсен, С. Б. Л., и Биф, Д. (2008). Механическое поведение стеклопластиковых композитных уплотнений для ИТ-ТОТЭ. Adv. Твердооксидные топливные элементы II Ceram. Англ. Sci . Proc . 27, 315–323. DOI: 10.1002 / 97804702
.ch41
CrossRef Полный текст | Google Scholar
Quadakkers, W. J., Piron-Abellan, J., Shemet, V., and Singheiser, L. (2003). Металлические соединители для твердооксидных топливных элементов — обзор. Mater. Высокий темп. 20, 115–127.DOI: 10.1179 / mht.2003.015
CrossRef Полный текст | Google Scholar
Родригес-Лопес, С. (2016). Thermomecánicas Propiedades de sellos vitrocerámicos системы RO-MgO-B2O3-SiO2 (R = Ba, Sr) для SOFC (кандидатская диссертация). CSIC, Мадридский автономный университет, Мадрид, Испания.
(2016). Thermomecánicas Propiedades de sellos vitrocerámicos системы RO-MgO-B2O3-SiO2 (R = Ba, Sr) для SOFC (кандидатская диссертация). CSIC, Мадридский автономный университет, Мадрид, Испания.
Google Scholar
Родригес-Лопес, С., Хаанапель, В. А. К., Дуран, А., Муньос, Ф., Мазер, Г. К., Паскуаль, М. Дж. И др. (2016). Стеклокерамические уплотнения в системе MgO-BaO-B 2 O 3 -SiO 2 Работа в смоделированных условиях. Внутр. J. Hydrog. Энергия 41, 15335–15345. DOI: 10.1016 / j.ijhydene.2016.07.051
CrossRef Полный текст | Google Scholar
Родригес-Лопес, С., Вей, Дж., Лауренти, К. К., Матиас, И., Хусто, В. М., Сербена, Ф. К. и др. (2017). Механические свойства стеклокерамических герметиков твердооксидных топливных элементов в системе BaO / SrO-MgO-B 2 O 3 -SiO 2 . J. Eur. Ceram. Soc. 37, 3579–3594. DOI: 10.1016 / j.jeurceramsoc.2017.03.054
CrossRef Полный текст | Google Scholar
Смеачетто, Ф., Chrysanthou, A., Moskalewicz, T., and Salvo, M. (2013). Термоциклирование соединенных конструкций Crofer22APU-герметик-анод с электролитом для планарных ТОТЭ до 3000 ч. Mater. Lett . 111, 143–146. DOI: 10.1016 / j.matlet.2013.08.025
CrossRef Полный текст | Google Scholar
Смеацетто, Ф., Хрисанту, А., Сальво, М., Чжан, З., и Феррарис, М. (2009). Характеристики и испытания стеклокерамического герметика, используемого для соединения электролита на анодной подложке с Crofer22APU в плоских твердооксидных топливных элементах. J. Источники энергии 190, 402–407. DOI: 10.1016 / j.jpowsour.2009.01.042
CrossRef Полный текст | Google Scholar
Смеачетто, Ф., Сальво, М., Феррарис, М., Казаленьо, В., Асинари, П., и Хрисанту, А. (2008). Характеристики и характеристики стеклокерамического герметика для соединения металлических межсоединений с YSZ и анодным электролитом в планарных ТОТЭ. J. Eur. Ceram. Soc . 28, 2521–2527. DOI: 10.1016 / j.jeurceramsoc.2008.03.035
J. Eur. Ceram. Soc . 28, 2521–2527. DOI: 10.1016 / j.jeurceramsoc.2008.03.035
CrossRef Полный текст | Google Scholar
Steinberger-Wilckens, R., Блюм, Л., Бухкремер, Х. П., де Хаарт, Б., Мальцбендер, Дж., И Пап, М. (2011). Последние результаты в разработке твердооксидных топливных элементов в Forschungszentrum Juelich. ECS Trans . 35, 53–60. DOI: 10,1149 / 1,3569978
CrossRef Полный текст | Google Scholar
Steinberger-Wilckens, R., Blum, L., Buchkremer, H.P., Gross, S., Bert de Haart, L., Hilpert, K., et al. (2006). Обзор разработки твердооксидных топливных элементов в Forschungszentrum Juelich. Внутр. J. Appl.Ceram. Технол . 3, 470–476. DOI: 10.1111 / j.1744-7402.2006.02102.x
CrossRef Полный текст | Google Scholar
Стивенс, Э. В., Ветрано, Дж. С., Коппель, Б. Дж., Чжоу, Ю., Сан, X. и Халил, М. А. (2009). Экспериментальная характеристика свойств стеклокерамических уплотнений и их реализация в моделях батарей твердооксидных топливных элементов. J. Источники энергии 193, 625–631. DOI: 10.1016 / j.jpowsour.2009.02.080
CrossRef Полный текст | Google Scholar
ThyssenKrupp VDM Gmb, H.(2010). Crofer22APU Лист технических данных № . 4046 (Вердохол).
Google Scholar
Тимуркутлюк Б., Цифлик Ю. и Коркмаз Х. (2015). Оценка прочности стеклокерамических композитов, содержащих оксид циркония, стабилизированный оксидом иттрия, после термоциклирования. Ceram. Инт . 41, 6985–6990. DOI: 10.1016 / j.ceramint.2015.01.153
CrossRef Полный текст | Google Scholar
Вэй, Дж., Пеканак, Дж., И Мальцбендер, Дж. (2015). Механические свойства стеклокерамических герметиков, армированных серебром, для твердооксидных топливных элементов. Ceram. Инт . 41, 15122–15127. DOI: 10.1016 / j.ceramint.2015.08.084
CrossRef Полный текст | Google Scholar
Вейль, К. С. (2006). Новейшие технологии герметизации твердооксидных топливных элементов. JOM 58, 37–44. DOI: 10.1007 / s11837-006-0052-6
JOM 58, 37–44. DOI: 10.1007 / s11837-006-0052-6
CrossRef Полный текст | Google Scholar
Вейл, К. С., Ким, Дж. Ю., и Харди, Дж. С. (2005). Реактивная пайка на воздухе: новый метод герметизации ТОТЭ и других твердотельных электрохимических устройств. Electrochem. Твердотельное Письмо . 8, A133 – A136. DOI: 10.1149 / 1.1850391
CrossRef Полный текст | Google Scholar
Янг, З., Харди, Дж. С., Уокер, М. С., Ся, Г., Симнер, С. П., и Стивенсон, Дж. У. (2004). Структура и проводимость термически выращенных окалин на ферритной Fe-Cr-Mn стали для соединений SOFC. J. Electrochem. Soc . 151: A1825. DOI: 10.1149 / 1.1797031
CrossRef Полный текст | Google Scholar
Ян З., Ся, Г., Мейнхард, К. Д., Вейл, К. С., Стивенсон, Дж. У. и Скотт, К. (2003). Химическая стабильность поверхностей раздела стеклянных уплотнений в твердооксидных топливных элементах с промежуточной температурой. J. Mater. Англ. Выполните . 22, 2892–2899. DOI: 10.1007 / s11665-013-0731-8
CrossRef Полный текст | Google Scholar
Чжао Ю. (2013) Термомеханические свойства стеклокерамических твердых оксидных герметиков для топливных элементов . Юлих: Forschungszentrum Jülich, RWTH Aachen University.
Google Scholar
границ | Внеклеточный матрикс скелетных мышц — что мы знаем о его составе, регуляции и физиологической роли? Повествовательный обзор
Введение
Скелетные мышцы являются важным компонентом состава тела человека, обычно составляя более 40% и 30% общей массы тела у мужчин и женщин, соответственно (Kim et al., 2002). Наиболее очевидная функция скелетных мышц — генерировать силы, необходимые для поддержания вертикальной позы и движения.Однако скелетные мышцы также играют важную роль во многих других физиологических процессах, включая термогенез (Rowland et al., 2015), метаболизм (Baskin et al., 2015) и секрецию многочисленных пептидов для связи с другими тканями (Pedersen and Febbraio , 2012). Таким образом, укрепление и поддержание здоровья скелетных мышц имеет жизненно важное значение. Хотя в последние годы фармакологические имитаторы упражнений вызывают все больший научный интерес (Fan and Evans, 2017), именно физические упражнения по-прежнему считаются наиболее мощным и универсально применимым инструментом для этих целей.
Таким образом, укрепление и поддержание здоровья скелетных мышц имеет жизненно важное значение. Хотя в последние годы фармакологические имитаторы упражнений вызывают все больший научный интерес (Fan and Evans, 2017), именно физические упражнения по-прежнему считаются наиболее мощным и универсально применимым инструментом для этих целей.
За последние десятилетия были проведены тысячи тренировочных исследований в попытке определить методы упражнений, наиболее подходящие для увеличения размера мышц и улучшения их функциональных характеристик в различных когортах (например, на момент написания этой рукописи Pubmed дал больше более 24 000 результатов по операторам поиска «упражнения» и «сила мышц»). Результаты этих исследований послужили источником вдохновения для различных рекомендаций по упражнениям, вероятно, наиболее известными из которых являются стенды с позициями, публикуемые и обновляемые нерегулярно Американским колледжем спортивной медицины (2009), Garber et al.(2011). Большинство исследований основывают свою оценку эффективности тренировочных вмешательств на исследовании сократительных мышечных клеток. Часто изучаемые параметры включают размер мышц, измеренный на уровне органа (Fisher et al., 2011) или на клеточном уровне (Schoenfeld, 2010), распределение типов волокон (Adams et al., 1993), архитектуру (Aagaard et al., 2001) как а также нервный импульс к мышцам (Folland and Williams, 2007).
Несмотря на обилие информации о пластичности скелетных мышц, часто упускается из виду тот факт, что мышечные волокна встроены во внеклеточный матрикс (ЕСМ), состоящий из сетки коллагеновых компонентов, а также смеси других макромолекул, таких как различные гликопротеины и протеогликаны.Недавние исследования показали, что ECM играет важную роль в развитии (Thorsteinsdóttir et al., 2011), росте (Fry et al., 2017) и восстановлении мышц (Calve et al., 2010), а также передаче сократительная сила (Street, 1983). Несмотря на то, что доказательства, демонстрирующие пластичность ЕСМ, существуют, лишь в небольшом количестве исследований сообщается о его реакциях на различные формы тренировок, предполагая, что физиологическая роль ЕСМ еще не полностью оценена специалистами по физическим упражнениям. Стремясь стимулировать дальнейшие исследования тренировочных реакций неконтрактильных компонентов скелетных мышц, мы предлагаем обзор текущего состояния знаний о составе, структуре и регуляции внеклеточного матрикса, его физиологической роли, нарушениях регуляции, связанных со старением и метаболизмом. расстройства, а также адаптации к физическим нагрузкам.
Стремясь стимулировать дальнейшие исследования тренировочных реакций неконтрактильных компонентов скелетных мышц, мы предлагаем обзор текущего состояния знаний о составе, структуре и регуляции внеклеточного матрикса, его физиологической роли, нарушениях регуляции, связанных со старением и метаболизмом. расстройства, а также адаптации к физическим нагрузкам.
Состав и структура скелетных мышц ECM
ECM скелетных мышц представляет собой сложную сеть, состоящую из коллагенов, гликопротеинов, протеогликанов и эластина (Takala and Virtanen, 2000; Halper and Kjaer, 2014).Коллагены образуют сеть внутримышечной соединительной ткани (IMCT), то есть центральные фиброзные компоненты ECM. IMCT обычно изображается организованной в виде трех слоев: (i) эндомизий, представляющий самый внутренний слой, который охватывает отдельные мышечные волокна, (ii) перимизий, связывающий группы мышечных волокон, и (iii) эпимизий, охватывающий всю мышцу. Большая структурная сложность сети ММКИ, подтвержденная сканирующими электронными микрофотографиями, предполагает, что эта традиционная классификация может быть упрощенной и что организация ЕСМ более высокого порядка еще нуждается в определении (Gillies and Lieber, 2011).Исследования фасциальных тканей также рассматривают слои IMCT как часть сложной системы взаимосвязанных и переплетенных соединительных тканей, которая «окружает, переплетается между собой и проникает во все органы, мышцы, кости и нервные волокна, обеспечивая тело функциональной структурой. и обеспечение среды, которая позволяет всем системам организма работать интегрированно » (Adstrum et al., 2017; Stecco et al., 2018). Эта система, которую обычно называют фасциальной системой, все чаще признается важной мишенью в спортивной медицине (Zügel et al., 2018).
IMCT содержит различные формы коллагенов, среди которых наиболее распространены типы I и III (Duance et al., 1977; Light and Champion, 1984; Gillies and Lieber, 2011; McKee et al., 2019). Эндомизий взаимодействует с сарколеммой миофибр на специальной базальной мембране, которая состоит в основном из коллагена IV типа и ламинина (Sanes, 1982; Martin and Timpl, 1987; Kjaer, 2004). Было обнаружено, что концентрация этих двух компонентов различается в зависимости от типа мышечных волокон, при этом медленно сокращающиеся волокна характеризуются значительно более высокими концентрациями коллагена IV, но более низкими концентрациями ламинина (Kovanen et al., 1988). Ламинин, в свою очередь, служит лигандом для двух сарколеммальных рецепторов — гликопротеинового комплекса, ассоциированного с дистрофином и интегрина α7β1 (Grounds et al., 2005), — расположенных на костамерах, которые представляют собой мембранные белковые структуры, выровненные в соответствии с Z диски миофибрилл. Считается, что интегрины действуют двунаправленным образом, позволяя внутриклеточным сигнальным молекулам регулировать внешнюю адгезию (передача сигналов «Inside-Out») и передавать внешние стимулы для воздействия на клеточные процессы (передача сигналов «Outside-In») (Boppart and Mahmassani, 2019) .Таким образом, явный стресс цитоскелета вызывает внутриклеточное связывание белков, таких как талин, винкулин или киндлин, что приводит к изменению конформации рецептора интегрина и позволяет внеклеточным доменам рецептора распространяться в направлении белков внутри ЕСМ. Кроме того, лиганды интегринов из внеклеточного пространства, такие как ламинин, коллаген или фибронектин, способствуют формированию высокоаффинного вертикального состояния, что приводит к усилению связывания с белками ЕСМ и кластеризации интегринов, особенно вдоль комплексов фокальной адгезии (Boppart and Mahmassani, 2019) .Гликопротеиновый комплекс, связанный с дистрофином, является еще одним важным фактором в обеспечении механической связи между сократительными компонентами скелетных мышц (т.е.актином) и взаимосвязанными слоями IMCT (Ervasti, 1993; Peter et al., 2011). Основные компоненты, связывающие сократительные элементы мышцы с интерстициальным матриксом, показаны на рисунке 1.
Рисунок 1. Основные компоненты внеклеточного матрикса скелетных мышц и его связь с сократительными компонентами мышцы.
Суперсемейство коллагена состоит в общей сложности из 28 различных членов, из которых типы I, III, IV, V, VI, XII, XIII, XIV, XV, XVIII и XXII, как было показано, присутствуют в зрелых скелетных мышцах в гене и / или уровень белка (Таблица 1). Формирующие фибриллы типы I и III, безусловно, наиболее многочисленны, и протеомные исследования показывают, что они вместе составляют примерно 75% от общего мышечного коллагена (McKee et al., 2019). Предполагается, что сильные параллельные волокна коллагена I типа, присутствующие в эндо-, пери- и эпимизиуме, придают мышцам прочность на разрыв и жесткость, тогда как коллаген III типа образует рыхлую сеть волокон, которая придает эластичность мышцам. эндо- и перимизий (Кованен, 2002).Коллаген типа IV, спиральная молекула, образует сетчатую структуру, которая составляет основу базальной пластинки (Sanes, 2003). Коллаген типа VI был обнаружен в эпимизиальном, перимизиальном и эндомизиальном интерстиции, но, в частности, в окрестностях базальной мембраны, где он взаимодействует с карбоксиконцевым глобулярным доменом коллагена типа IV (Kuo et al., 1997). Интересно, что коллаген VI имеет нетипичные неколлагеновые участки, образующие отчетливую микрофибриллярную сеть в большинстве соединительных тканей (Maaß et al., 2016). Мутации коллагена VI приводят к заболеваниям с сочетанным вовлечением мышц и соединительной ткани, включая врожденную мышечную дистрофию Ульриха, миопатию Бетлема, аутосомно-доминантную мышечную дистрофию пояса конечностей и аутосомно-рецессивный миосклероз (Bushby et al., 2014).
Таблица 1. Обзор коллагеновых компонентов внеклеточного матрикса скелетных мышц.
Коллаген типов XII, XIV, XIX и XXII относится к коллагенам, ассоциированным с фибриллами, с прерывистыми тройными спиралями (FACIT; Chiquet et al., 2014; Calvo et al., 2020), посредством чего коллаген типа XXII, по-видимому, экспрессируется исключительно в тканевых соединениях, таких как мышечно-сухожильные соединения в скелетных и сердечных мышцах (Koch et al., 2004).
Биоинформатические инструменты для скрининга протеома человека нормальных и больных тканей позволили охарактеризовать глобальный состав протеома ЕСМ, или «матрисомы». В общей сложности 1027 генов были связаны с ЕСМ, посредством чего основные белки матрисомы (гликопротеины ЕСМ, коллагены и протеогликаны) можно было отличить от белков, связанных с матрисомами (аффилированные с ЕСМ белки, регуляторы ЕСМ и секретируемые факторы, которые могут взаимодействовать с основными белками). Белки ЕСМ) (Naba et al., 2016). Учитывая сложность ткани скелетных мышц человека, включающей многоядерные мышечные волокна, иммунные клетки, эндотелиальные клетки, мышечные стволовые клетки, немиогенные мезенхимальные предшественники и другие мононуклеарные клетки (Bentzinger et al., 2013a), для выяснения этого потребуются будущие исследования. вклад каждой из этих клеток в структуру и реконструкцию IMCT. Сигнатуры генов, полученные, например, из последовательности РНК изолированных мышечных волокон и других типов клеток, представляют собой многообещающий инструмент для деконволюции основной массы скелетной мышечной ткани (Rubenstein et al., 2020).
Физиологическая регуляция генов ЕСМ
Гомеостаз ЕСМ поддерживается посредством точно настроенных анаболических и катаболических процессов, которые регулируются различными факторами роста, протеогликанами и ферментами, ответственными за деградацию коллагена. Было обнаружено, что после связывания с мембранно-связанными рецепторами факторы роста, принадлежащие к суперсемейству трансформирующих факторов роста бета (TGF-β), индуцируют фосфорилирование белков Smad, которые передают внеклеточные сигналы в ядро, где они активируют транскрипцию нижележащих генов, что приводит к продукции коллагена. (Макдональд и Кон, 2012).Другим, хотя и менее описанным, фактором аналогичной функции является фактор роста соединительной ткани (CTGF), избыточная экспрессия которого, как сообщается, вызывает дистрофический фиброз мышц и функциональные нарушения (Morales et al., 2011).
Функция этих анаболических факторов в основном регулируется небольшими протеогликанами, богатыми лейцином (SLRP). Декорин, прототип этого семейства, деактивирует профибротический TGF-β и CTGF (Zhu et al., 2007; Brandan and Gutierrez, 2013), а также ограничивает фибриллогенез путем прямого связывания с коллагеном I типа (Reese et al., 2013). Другой SLRP представлен бигликаном, который конкурирует с декорином за тот же сайт связывания на коллагене (Schönherr et al., 1995) и, вероятно, играет роль как в формировании, так и в регенерации мышц (Brandan et al., 2008).
Транскрипционная регуляция образования белков, по-видимому, является важным фактором пластичности ECM. В этом отношении было показано, что экспрессия белка в скелетных мышцах слабо регулируется на уровне мРНК, что приводит к большим различиям в содержании мРНК и белка в различных тканях (Wang et al., 2019). Интересно, что характер регуляции белка зависит от функции белка, посредством чего ассоциация между мРНК и белком выше для ECM и организации коллагеновых фибрилл (Makhnovskii et al., 2020). Другим интересным аспектом регуляции количества белков ЕСМ является тот факт, что индукция транскрипции, по-видимому, для коллагена довольно медленная, поскольку для полной индукции транскрипции требуется почти 3 дня. Напротив, скорость секреции адаптируется быстро, поскольку она повышается менее чем за 1 час.В клетках с высоким уровнем продуцирования коллагена этот путь контролируется посттранскрипционной регуляцией, которая требует контроля с обратной связью между скоростями секреции и трансляции (обзор у Schwarz, 2015).
Что касается ремоделирования ткани, два семейства ферментов, матриксные металлопротеиназы (MMP) и тканевые ингибиторы металлопротеиназ (TIMP), участвуют в регуляции гомеостаза ECM. ММП представляют собой протеолитические ферменты, которые разрушают различные типы коллагенов и ингибируются ТИМП (Visse and Nagase, 2003; Alameddine, 2012).В частности, MMP-1 и MMP-8 инициируют деградацию коллагенов I и III (преобладающих в эндо-, пери- и эпимизиуме), тогда как MMP-2 и MMP-9 расщепляют коллаген IV типа (основной коллагеновый компонент базальная мембрана) (Corcoran et al., 1996). ТИМП-1, -2 и -4 способны ингибировать все известные ММП (Christensen and Purslow, 2016).
ECM и сила скелетных мышц
Взаимодействие актина и миозина, а также многих других саркомерных белков приводит к укорочению мышечных волокон.Традиционные биомеханические модели часто изображают мышечно-сухожильные единицы как системы, в которых силы, возникающие в результате укорочения волокна, передаются продольно вдоль мышечного волокна и далее, в мышечно-сухожильном соединении, на сухожилие. Близко к мышечно-сухожильному соединению миофибры имеют пальцевидные отростки, которые образованы из инвагинаций плазматической мембраны (Knudsen et al., 2015). Эта структура увеличивает площадь поверхности, доступную для передачи усилия. Ожидается, что передача силы будет происходить между пальцевидными отростками мышечного волокна и коллагеновыми волокнами, расположенными внутри инвагинаций, посредством срезания базальной пластинки (Huijing, 1999).Коллагены, содержащиеся здесь, относятся к типу XXII, который образует внутренний слой, а также к III, VI, XII и XIV, которые расположены дальше от мембраны мышечных волокон (Jakobsen et al., 2017). Хотя его точная роль до сих пор неясна, интересно отметить, что в мышцах коллаген XXII располагается исключительно в мышечно-сухожильном соединении. У рыб-зебр дефицит коллагена XXII, как было обнаружено, приводит к мышечной дистрофии (Charvet et al., 2013), предполагая, что этот коллаген может служить для поддержания структурной целостности и стабилизации мышечно-сухожильного соединения.
Однако, учитывая тот факт, что значительная часть волокон в длинных мышцах оканчивается внутрипучко, не достигая непосредственно сухожилия (Barrett, 1962; Hijikata et al., 1993), становится ясно, что мышечно-сухожильный путь не может представлять собой единственный механизм передачи силы. . Intrafascicularly согласующего волокно должно полагаться на носителе, расположенный параллельно с ними, чтобы передать свои силы на пассивных компоненты опорно-двигательной системы (Sheard, 2000). Как впервые было обнаружено Стрит (1983), именно сеть ММКИ внутри ВЦМ способствует такой боковой передаче сократительной силы.Передача силы через сеть IMCT происходит от сократительных белков через костамеры к эндомизию (Bloch and Gonzalez-Serratos, 2003; Peter et al., 2011) — как показывают исследования моделирования, посредством сдвига (Sharafi and Blemker, 2011; Zhang and Gao, 2012). ) — и далее в перимизий, который окончательно сливается с апоневрозами и сухожилиями (Passerieux et al., 2007). Первые сведения о пропорциях передачи продольной и поперечной силы в поперечно-полосатых мышцах получены из элегантных экспериментов Huijing et al.(1998). После разрыва прямых соединений нескольких головок длинного разгибателя пальцев рук крысы, соответствующих 55% общей мышечной массы, от суставного сухожилия, Huijing et al. (1998) наблюдали, что сила поддерживалась на уровне 84% от силы неповрежденной мышцы. Совсем недавно Ramaswamy et al. (2011) использовали коромысло для прямого измерения сил, передаваемых по продольным и поперечным путям. Их результаты не только подтвердили, что более 50% силы передавалось в боковом направлении, но также показали, что передача боковой силы была значительно снижена как у дистрофических, так и у старых грызунов.Позже их результаты были подтверждены Чжаном и Гао (2014).
Некоторые аргументы предполагают, что боковая передача силы является биомеханической необходимостью для поддержания целостности мышц и повышения эффективности сокращения. Во-первых, он помогает распределять сократительные силы по всей поверхности миофибрилл, что снижает механическое напряжение и защищает волокна от чрезмерного растяжения. Это может быть особенно важно в концевых областях волокон, которые обычно имеют конус и поэтому не подходят для выдерживания чрезмерных усилий (Monti et al., 1999). Косвенная поддержка этой гипотезы обеспечивается исследованиями с участием пожилых людей (Hughes et al., 2016) или пациентов, страдающих дистрофией Дюшенна (Virgilio et al., 2015), у которых дистрофин (т.е. костамерный белок, устанавливающий механическую связь между цитоскелет, сарколемма и ECM и, таким образом, способствует передаче латеральной силы) либо утрачены, либо нарушены, а восприимчивость к травмам от растяжения мышц увеличивается.
Кроме того, считается, что передача поперечной силы приводит к сокращению волокон моста либо в разное время, либо в неравной степени (Yucesoy et al., 2006), что поможет поддерживать выравнивание волокон и, таким образом, структурную целостность мышцы (Purslow, 2002). Недавно Dieterich et al. (2017) сравнили начало сокращения, определенное электромиографией и УЗИ в M-режиме. Как ни странно, авторы обнаружили, что начало движения предшествует электромиографическому сигналу в ~ 20% исследований, что можно объяснить переносом латеральной силы. Действительно, хотя продольная передача сил может задерживаться из-за необходимости натягивать эластичные элементы, размещенные последовательно с мышцей (Nordez et al., 2009), трансламинарное сцепление сдвига между мышечными волокнами и сетью IMCT может обеспечить немедленную передачу силы. Наконец, передача боковой силы обеспечивает механизм, посредством которого сила может по-прежнему генерироваться и передаваться от мышечных волокон, которые прерваны из-за микротравм или во время роста мышц (Purslow, 2010).
Помимо своей роли в латеральном переносе сократительной силы, ECM может также влиять на укорочение мышечных волокон. Часто предполагается, что сократимость миофибрилл ограничивается геометрией составляющих его саркомеров: саркомера и, таким образом, укорачивание волокна прекращается, когда полосы z входят в контакт с миозиновыми нитями.Однако эти идеи рассматривают только поведение саркомера как независимого исполнительного механизма. В условиях in vivo мышечные волокна встраиваются в сеть IMCT, что может препятствовать укорочению волокон. Действительно, принцип постоянного объема (Baskin and Paolini, 1967) диктует, что во время укорачивания мышечные волокна должны подвергаться радиальному расширению, что уже давно экспериментально подтверждено даже на саркомерном уровне (Brandt et al., 1967). Новые вычислительные модели и измерений in situ, измерений в мышцах лягушки Азизи и др.(2017) продемонстрировали, что укорачивание мышц затрудняется, когда радиальное расширение ограничивается физическими ограничениями. Следовательно, изменения количества и механических свойств сети IMCT, в которую встроены мышечные волокна, могут напрямую влиять на сократимость скелетных мышц. Такой сценарий может быть представлен мышечным фиброзом (Gillies et al., 2017).
ECM в развитии, росте и восстановлении скелетных мышц
Помимо передачи силы, ECM скелетных мышц выполняет несколько важных функциональных ролей.Очевидно, сеть ММКИ обеспечивает механическую поддержку мышечным волокнам, а также поддерживающим их нервам и кровеносным сосудам. Кровеносные капилляры проходят в промежутках, занятых эндомизием, их количество и плотность зависят от размера мышечных волокон (Janácek et al., 2009). Помимо этой наиболее очевидной роли, взаимодействие между миобластами, дифференцированными мышечными волокнами и компонентами внеклеточного матрикса имеет центральное значение для эмбриогенного развития, дальнейшего роста и восстановления мышечной ткани.
Клеточным источником коллагеновых компонентов мышечного ECM являются специальные фибробласты IMCT, которые происходят из различных эмбриогенных источников, включая сомиты (Nowicki et al., 2003), мезодерму латеральной пластинки (Pearse et al., 2007) и нервную систему. клетки гребня (Olsson et al., 2001). Поскольку они продуцируют не только фибробласты, но и адипогенные клетки, фибробласты IMCT могут рассматриваться как фиброипогенные предшественники (Uezumi et al., 2010). Недавние исследования предоставили доказательства того, что, помимо этих очевидных ролей, фибробласты IMCT и производимые ими соединительные ткани влияют на миогенез (т.е.д., образование мышечных предшественников и их дифференциация в многоядерные миофибриллы) и морфогенез мышц (то есть процесс, в котором миофибры собираются в мышцы), таким образом действуя как важные регуляторы развития мышц. Эти сложные регуляторные процессы, происходящие во время эмбриогенного развития, здесь подробно не рассматриваются, но были подробно рассмотрены в других источниках (Nassari et al., 2017; Sefton and Kardon, 2019). Вкратце, IMCT направляет мышечных предшественников к их назначенным областям-мишеням посредством комбинации привлекательных (фактор роста гепатоцитов, фактор, производный стромальными клетками) и отталкивающих сигналов (эфрин) (Dietrich et al., 1999; Swartz et al., 2001). Посредством множества факторов транскрипции, экспрессируемых в фибробластах IMCT, IMCT затем способствует пролиферации, выживанию и дифференцировке соседних миобластов в зрелые миофибриллы (Kardon et al., 2003; Hasson et al., 2010; Iwata et al., 2013; Vallecillo -García et al., 2017). Таким образом, можно предположить, что IMCT служит мезодермальным препаттерном, который контролирует участки дифференцировки миофибрилл и, следовательно, окончательное положение, размер и форму мышц.
Будучи постмитотическими тканями, скелетные мышцы зависят от сателлитных клеток для адаптации и регенерации на протяжении всей жизни. Эти стволовые клетки располагаются в специализированных нишах между сарколеммой мышечных волокон и их инкапсулирующими базальными мембранами. Поддержание, активация и дифференцировка сателлитных клеток регулируются сложными каскадами факторов транскрипции. Для подробного обзора этих сотовых схем читатели могут обратиться к недавнему обзору Almada and Wagers (2016). Имея особое отношение к этой рукописи, растущее количество доказательств указывает на то, что на судьбу сателлитных клеток также сильно влияют взаимодействия с нишей ECM, в которой они находятся.Действительно, как динамическая среда ниша стволовых клеток передает механические и химические сигналы, которые действуют, защищая покоящиеся стволовые клетки или индуцируя активацию, пролиферацию и дифференцировку.
В состоянии покоя сателлитные клетки экспрессируют канонический клеточный регулятор спаренный бокс-белок 7 (PAX7) (Olguin and Olwin, 2004). Исследования in vitro показали, что большая часть сателлитных клеток экспрессирует PAX7 при культивировании на матригеле, смеси белков ЕСМ и факторов роста (Wilschut et al., 2010; Грефте и др., 2012). Дальнейшее подтверждение идеи, что ECM активно участвует в поддержании покоя сателлитных клеток, происходит из сообщений о том, что сателлитные клетки, удаленные из своей ниши, быстро входят в клеточный цикл и теряют свою способность к миогенной дифференцировке (Gilbert et al., 2010). Интересно, что сателлитные клетки, по-видимому, также способны воспринимать различные механические свойства ECM и реагировать на них. Фактически, экспрессия PAX7 и выживаемость сателлитных клеток выше при культивировании на гидрогелях, имитирующих физиологическую жесткость мышц (Gilbert et al., 2010). Кроме того, сателлитные клетки, культивируемые на мягком гидрогеле, обладают большей функциональной способностью после трансплантации в мышцу реципиента (Cosgrove et al., 2014).
Кроме того, было показано, что компоненты ECM влияют на деление стволовых клеток. В частности, белки фибронектин (Bentzinger et al., 2013b) и коллаген VI (Urciuolo et al., 2013), а также протеогликаны синдекан 3, синдекан 4, перлекан и декорин (Cornelison et al., 2001; Brack et al. ., 2008) были идентифицированы как составляющие ниши, влияющие на баланс между дифференцировкой и самообновлением и, таким образом, на поддержание регенеративной способности скелетных мышц.
При мышечной травме или в ответ на повышенную нагрузку обычно в основном покоящиеся сателлитные клетки активируются и дифференцируются в миобласты, чтобы окончательно слиться в зрелые миофибриллы. Хотя этот процесс требует своевременной экспрессии различных факторов транскрипции, таких как миогенный фактор 5, белок миогенной детерминации или миогенин (Almada and Wagers, 2016), несколько исследований указывают на влияние ECM на каждый из этих этапов. Эксперименты с миобластами мышей (Grefte et al., 2012) или свиней (Wilschut et al., 2010) показали, что на слияние миобластов положительно влияет матригель, но не отдельные субстраты, присутствующие в нише ECM. Вклад отдельных белков все еще плохо изучен, однако сопутствующее присутствие поли- D -лизина и ламинина (Boonen et al., 2009), гликозаминогликанов (Rønning et al., 2013) и протеогликанов сульфата гепарина (Gutiérrez и Brandan, 2010), по-видимому, играют важную роль в пролиферации и дифференцировке сателлитных клеток. После активации стволовых клеток скелетных мышц локальное ремоделирование ВКМ сопровождается отложением ламинина-α1 и ламинина-α5 в базальной пластинке ниши сателлитных клеток (Rayagiri et al., 2018). На мышах было показано, что мышечные сателлитные клетки продуцируют коллагены ЕСМ для поддержания покоя клеточно-автономным образом, при этом коллаген V является критическим компонентом покоящейся ниши, поскольку истощение приводит к аномальному входу в клеточный цикл и постепенному уменьшению стволовых клеток. бассейн (Багдади и др., 2018). Так же, как для поддержания покоя, адекватные механические свойства ниши ECM также могут быть важны для созревания сателлитных клеток. Действительно, было обнаружено, что миотрубки оптимально дифференцируются на субстратах с мышечной жесткостью (Engler et al., 2004). В совокупности эти данные предполагают, что усиление жесткости ВКМ, сопровождающее как различные скелетно-мышечные нарушения, так и процесс старения, может отрицательно влиять на регенеративную способность мышцы.
Ремоделирование мышц ECM с возрастом
В более старшем возрасте скелетные мышцы обычно демонстрируют фиброзную морфологию (Lieber and Ward, 2013). В отличие от фасциального уплотнения, при котором может сохраняться общая структура коллагенов (Pavan et al., 2014), возрастной мышечный фиброз характеризуется потерей четкой двунаправленной ориентации решетки здоровых перимизиальных волокон коллагена и его замещением. из-за неустойчивой оптоволоконной сети со сниженным образованием складок (Ярвинен и др., 2002). Кроме того, может увеличиваться абсолютное содержание коллагена и (неферментативное) сшивание коллагеновых волокон (Haus et al., 2007b). Таким образом, модуль упругости ЕСМ может быть увеличен примерно в 35 раз (с ~ 12 кПа в молодых до ~ 418 кПа в старых мышцах; Yin et al., 2013), причем этот эффект обусловлен накоплением плотно упакованных и сильно сшитый коллаген (Wood et al., 2014). У крупных и долгоживущих животных, таких как тюлени Уэдделла, наблюдалось увеличение внеклеточного пространства на 35-40% по мере увеличения общего и относительного содержания коллагена с возрастом.Однако это увеличение связано со сдвигом в сторону более высокого отношения коллагена типа I к коллагену типа III (Hindle et al., 2009). Более того, концентрация коллагена IV типа повышается в базальной пластинке медленно сокращающихся мышц, тогда как концентрация ламинина, по-видимому, снижается с возрастом (Kovanen et al., 1988). Было также показано, что повышенное отложение белков базальной пластинки вытесняет сателлитные клетки из их ниш, что влияет на регуляцию делений сателлитных клеток (Snow, 1977) и может объяснять меньшее количество сателлитных клеток, обычно учитываемых в старых мышцах по сравнению с молодыми мышцами. (Brack et al., 2007). Утрата и функциональная инактивация стволовых клеток, которые негативно влияют на гомеостаз тканей, могут рассматриваться как общий признак старения (López-Otín et al., 2013), который следует рассматривать как универсальную силу, приводящую к старению мышц (Brack and Muñoz-Cánoves, 2016) и другие ткани (Oh et al., 2014). Помимо воздействия на сателлитные клетки, нерегулируемая базальная пластинка также, как ожидается, нарушит регенеративную способность мышц из-за неадекватной поддержки мышечных волокон и дезорганизованной ориентации каркаса (Sanes, 2003).Обзор, включающий обширный обзор эффектов старения на ECM скелетных мышц, был недавно опубликован Etienne et al. (2020).
Интересно, что данные транскрипционного профилирования мышц молодых и старых крыс предполагают, что из 682 наборов зондов, которые значительно различались между молодыми и старыми животными, 347 генов фактически уменьшились (а не увеличились) в старых / саркопенических мышцах по сравнению с молодыми мышцами. Было показано, что из этих генов 24% играют биологическую роль в ECM и клеточной адгезии (Pattison et al., 2003). Эти данные подтверждают гипотезу, что возрастные изменения в ECM могут быть вызваны сниженной способностью к деградации, а не повышенным синтезом коллагеновых структур. В частности, ММП, по-видимому, играют важную роль в этих процессах (de Sousa Neto et al., 2018). Это также подтверждается данными, которые предполагают снижение способности структур ВКМ к ремоделированию структур ВКМ в старых мышцах, вызванное упражнениями с отягощениями (Wessner et al., 2019). Хотя механизмы еще полностью не изучены, также считается, что эти изменения напрямую ухудшают функцию мышц, препятствуя сократимости волокон (Azizi et al., 2017) и передача поперечной силы (Sharafi and Blemker, 2011).
Нарушение регуляции ECM скелетных мышц вследствие метаболических нарушений
Хорошо известно, что скелетные мышцы играют важную роль в стимулированном инсулином поглощении глюкозы (Richter and Hargreaves, 2013). Роль ECM в этом контексте может быть менее ясной. Мышцы с дефицитом β1-специфического интегрина демонстрируют снижение скорости инфузии глюкозы, стимулированной инсулином, и клиренса глюкозы, несмотря на отсутствие изменений в потреблении пищи, весе, глюкозе натощак, уровнях инсулина и экспрессии белка GLUT4 (Zong et al., 2009), подтверждая связь между аберрантной передачей сигналов интегрина и развитием диабета 2 типа. Более того, на животной модели диабета было показано, что нарушение чувствительности к инсулину связано со снижением уровней белка изоформы дистрофина Dp427 и субкомплекса альфа / бета-дистрогликана (Mulvey et al., 2005).
Повышенное количество коллагена типа I и III было обнаружено как у пациентов с диабетом 2 типа, так и у недиабетиков с ожирением (Berria et al., 2006), а перекорм у людей был связан с увеличением экспрессии генов, связанных с IMCT (коллагены I, III, IV, V, SPARC, интегрин; Tam et al., 2014) и изменения генных путей, связанные с взаимодействием рецепторов ECM, фокальной адгезией и сращиванием (Tam et al., 2017). Однако кормление мышей диетой с высоким содержанием жиров привело к снижению уровней экспрессии генов COL1, COL3 и COL6, но не уровня белка (Tam et al., 2015).
Деградация коллагенов посредством MMPs, как было показано, является важным компонентом ремоделирования ECM (Cui et al., 2017). Может ли это быть правдой и в контексте диабета, было исследовано в исследовании на животных.Интересно, что генетическое истощение MMP9 не вызывало инсулинорезистентности у худых мышей, несмотря на то, что приводило к увеличению коллагена IV. Однако, когда мышей кормили диетой с высоким содержанием жиров, делеция вызывала глубокое состояние инсулинорезистентности. Эти результаты еще больше усиливают роль компонентов ММКИ в развитии мышечной инсулинорезистентности, особенно в состоянии переедания (Kang et al., 2014).
Наконец, гиалуронан, основной компонент ЕСМ, увеличивается при ожирении у мышей, вызванном диетой с высоким содержанием жиров.Для лечения инсулинорезистентности с сопутствующим уменьшением жировой массы, размера адипоцитов, а также печеночной и мышечной инсулинорезистентности (Kang et al., 2013) предлагается лечение с помощью PEGPh30, который дозозависимо снижает гиалуронан в мышечном ECM.
Подводя итог, можно сказать, что различные компоненты ECM подвержены влиянию на различных стадиях диабета. Исследования о том, связан ли диабет со слабостью мышц, противоречивы (Leong et al., 2015; Li et al., 2016), и еще предстоит выяснить, имеют ли непосредственное отношение к этому контексту изменения в проводящих путях, связанных с ECM.
Адаптация к физическому воспитанию и неиспользование
Первые свидетельства, указывающие на гибкость ММКИ в ответ на физическую активность, были опубликованы еще в 1970-х годах, когда Suominen и Heikkinen (1975) и Suominen et al. (1977) обнаружили более высокие уровни пролилгидроксилазы (фермента, способствующего биосинтезу коллагена) у тренированных на выносливость спортсменов, а также в продольном исследовании после восьми недель аэробных тренировок. Позже было обнаружено, что влияние упражнений на выносливость на активность проколлагеновых ферментов более выражено у красных мышц, чем у белых (Takala et al., 1983). Прямые измерения содержания коллагена, впервые проведенные в конце 1980-х годов, подтвердили, что содержание коллагена (типа IV) увеличилось в устойчивой к усталости камбаловидной мышце крыс после пожизненных тренировок на выносливость (Kovanen et al., 1988). Несмотря на вызванное физическими упражнениями увеличение коллагена, Gosselin et al. (1998) обнаружили, что укреплению мышц, наблюдаемому с возрастом, можно противодействовать с помощью упражнений на выносливость, которые авторы связывают с уменьшением сшивания гидроксилизилпиридинолином коллагеновых волокон.
Эффекты иммобилизации на ECM скелетных мышц не совсем однозначны. Ранние исследования Karpakka et al. (1990, 1991) обнаружили, что у крыс снижается как активность гидроксилазы, так и содержание гидроксипролина (аминокислоты, составляющей коллагены). Позже было обнаружено, что изменения содержания коллагена в ответ на кратковременную иммобилизацию или неиспользование были довольно небольшими (Savolainen et al., 1988; Haus et al., 2007a), что можно объяснить относительно низкой скоростью обновления. В более позднем исследовании, напротив, было обнаружено, что содержание коллагена I и биомеханические свойства (модуль упругости, максимальное напряжение и предел текучести) фасции голени, покрывающей трехглавую мышцу surae крысы, значительно увеличиваются после всего лишь 21 дня разгрузки задней конечности. (Хуанг и др., 2018). Интересно, что эти изменения можно предотвратить, применяя вибрацию к задним лапам крыс дважды в день. У людей, не занимающихся спортом, иммуногистохимическое окрашивание не показало изменений в плотности сети коллагена I после 60 дней постельного режима. Напротив, у субъектов, выполняющих протокол контрмер, состоящий из реактивных прыжков на санях, иммунореактивность коллагена I была снижена по сравнению с исходными уровнями (Schoenrock et al., 2018).
Еще одна модель, которая позволяет изучить адаптивность ECM мышц, — это функциональная перегрузка, вызванная устранением хирургического синергиста.В одном из первых соответствующих исследований Уильямс и Голдспинк (1981) перерезали сухожилия подошвенных и икроножных мышц самцов крыс, чтобы перегрузить камбаловидные мышцы. Гипертрофия мышц, наблюдаемая через три недели после тенотомии, сопровождалась увеличением концентрации IMCT (> 45%) и отношения IMCT к мышечной ткани. Гистологический анализ также показал, что увеличение IMCT в основном связано с утолщением эндомизия. Сосредоточившись на мышечно-сухожильном соединении, Zamora и Marini (1988) провели аналогичные эксперименты и изолировали подошвенную мышцу крысы с помощью тенотомии камбаловидной мышцы и удаления икроножных мышц.По сравнению с контрольными животными фибробласты, расположенные в мышечно-сухожильном соединении, развивали более высокую степень активации цитоплазмы, ядра и ядрышка уже после одной-двух недель функциональной перегрузки. В более недавнем исследовании было проверено влияние ИЛ-6 на ремоделирование ВКМ, вызванное перегрузкой, путем сравнения мышей дикого типа и мышей с нокаутом по ИЛ-6 (White et al., 2009). В то время как увеличение площади поперечного сечения миофибрилл было одинаковым после 21 дня функциональной перегрузки, увеличение сырого веса мышц было значительно больше у мышей с нокаутом по IL-6.Гистологический анализ подтвердил, что этот избыточный прирост мышечной массы можно объяснить значительно большим увеличением содержания несократительной ткани и концентрации гидроксипролина, что свидетельствует о содержании коллагена и фиброзе. В соответствии с этим наблюдением, уровни мРНК проколлагена-1, IGF-1 и TGF-β были значительно выше у мышей с дефицитом IL-6. Напротив, экспрессия мРНК MyoD, фактора транскрипции, необходимого для мио-, а не фиброгенной дифференцировки сателлитных клеток (Zammit, 2017), была значительно ослаблена у животных, лишенных IL-6.В совокупности эти результаты показывают, что устранение синергиста вызывает увеличение содержания IMCT и, в частности, утолщение эндомизиальных структур в перегруженных мышцах. Эти адаптации могут служить для модуляции несокращающихся структур мышц в соответствии с повышенными функциональными требованиями. IGF-1, по-видимому, играет важную роль в регуляции этого процесса, поскольку было показано, что недостаток IGF-1 приводит к чрезмерному накоплению IMCT и, возможно, к нарушению регенеративного потенциала мышц.
Одно из первых исследований по тестированию и сравнению различных форм упражнений с отягощениями у мужчин было проведено Brown et al.(1999), которые сообщили, что после одного приступа концентрических сокращений маркеры распада коллагена (гидроксипролин и коллаген сывороточного типа I) не увеличивались. Напротив, эксцентрическое действие мышц увеличивало уровень коллагена в сыворотке более чем на 40% в течение 9 дней после тренировки, что указывает на то, что эксцентрические сокращения могут быть более эффективными в стимулировании распада коллагена. Эти результаты были подтверждены в двух более поздних исследованиях, аналогичным образом использовавшихся с использованием высокоинтенсивных эксцентрических упражнений, которые выявили как повышенный процессинг проколлагена и содержание коллагена типа IV, так и более высокую активность ММП и ТИМП (Crameri et al., 2004; Mackey et al., 2004). Интересно, что Crameri et al. (2004, 2007) также сообщили об увеличении тенасцина С, гликопротеина, присутствующего в ЕСМ, который, как предполагается, направляет миграцию клеток после повреждения, независимо от того, было ли повреждение мышц вызвано произвольным или электрически индуцированным повреждением мышц. Временное повышение активности тенасцина С и других гликопротеинов ЕСМ (например, фибронектина и гиалуроновой кислоты) обычно называют «переходной матрицей», появление которой считается важным первым шагом для успешного восстановления мышц, так как дает важные сигналы, управляющие регенеративный потенциал мышечных стволовых клеток (Calve et al., 2010; Tierney et al., 2016). Сообщается, что высвобождение гликопротеинов ECM сопровождается повышенной активностью MMP-9 у молодых, но снижением активности MMP-9 и MMP-15 у пожилых людей (Wessner et al., 2019). Эти данные свидетельствуют о том, что острые упражнения с отягощениями вызывают катаболический ответ в молодых мышцах, но этот эффект может быть нарушен в более старшем возрасте. Последующая анаболическая реакция, характеризующаяся повышенной регуляцией структурных коллагенов (I, III, IV) и ламинина, как было обнаружено, происходит со значительной задержкой, что позволяет предположить, что восстановление мышц, вызванное острым приступом повреждающих (удлинительных) сокращений, следует за двухфазный характер (Mackey et al., 2011; Hyldahl et al., 2015). Интересно, что недавнее исследование Sorensen et al. (2018) обнаружили, что внешний вид переходной матрицы у физически активных пожилых людей притупляется по сравнению с молодыми людьми. Это наблюдение поддерживает представление о том, что дисрегулируемые сигналы ECM могут быть ответственны за увеличение отложения ECM и снижение активности стволовых клеток, обычно наблюдаемых в более старых мышцах (Grounds, 1998).
Одно из первых исследований по прямому сравнению различных форм мышечных сокращений с точки зрения их острого ремоделирующего потенциала ВКМ было опубликовано Heinemeier et al.(2007). Эти авторы провели исследование на грызунах и обнаружили, что активность генов, связанных с биосинтезом коллагена (например, коллагенов I и III), а также факторов роста (например, TGF-β1) повышается после всех форм физических упражнений, но наиболее заметно. после эксцентрической тренировки. В организме человека, коллаген белок дробные скорости синтеза также были предложены, чтобы быть более возросли после острого приступа эксцентричных по сравнению с концентрическим обучением (Холм и др., 2017), хотя это понятие не неоспоримое (Moore и др., 2005). В совокупности эти результаты предполагают, что особенно эксцентрические упражнения являются мощным стимулом, который вызывает микротравмы и обмен клеток IMCT, причем последний, как предполагается, представляет собой попытку организма предотвратить повторное повреждение мышцы (Mackey et al., 2011; Hyldahl et al. , 2015; Takagi et al., 2016). Фактически, снижение активности ММП после продолжительной тренировки, состоящей из электрически вызванных изометрических сокращений у крыс, может отражать успешное усиление ВКМ (Ogasawara et al., 2014), тогда как продолжительное повышение активности ММП и ТИМП в подошвенной мышце мышей после хирургического удаления икроножной и камбаловидной мышц может указывать на продолжающееся ремоделирование ВКМ (Mendias et al., 2017).
Помимо режима сокращения, ECM скелетных мышц также может быть чувствительным к интенсивности упражнений. Carmeli et al. (2005) протестировали влияние бега на беговой дорожке с высокой или низкой интенсивностью на крысах и обнаружили, что MMP-2 (один из ферментов, ответственных за расщепление коллагена IV, в основном присутствующего в базальной мембране мышц) повышается после упражнений высокой интенсивности. Только. На людях, напротив, одно исследование Holm et al. (2010) сравнили эффекты односторонних упражнений на разгибание колен, выполняемых с низкой или высокой (16% или 70% от индивидуального максимума одного повторения, соответственно) интенсивностью, с количеством повторений, скорректированным в соответствии с вмешательствами для общей нагрузки. поднял.В этом исследовании скорость фракционного синтеза коллагена была равномерно увеличена после обоих вмешательств.
Что касается адаптации ЕСМ к длительным тренировкам с отягощениями, существуют только данные исследований на животных. де Соуза Нето и др. (2018) сообщили, что 12 недель тренировок с отягощениями, состоящих из подъемов по лестнице с прогрессивными дополнительными нагрузками, эквивалентными 65–100% максимальной несущей способности каждого индивидуума, активировали активность ММП-2 в подошвенных сгибательных мышцах старых крыс, в то же время подавляя ММП-2. 2 и ММП-9 в кровообращении.Вывод авторов о том, что тренировки с отягощениями могут, таким образом, быть полезным инструментом для поддержания ремоделирования ВКМ в более старшем возрасте, недавно получил эмпирическую поддержку в другом исследовании обучения на крысах, которое использовало тот же протокол обучения и показало пониженное отложение соединительной ткани у тренированных пожилых людей. мышцы (Guzzoni et al., 2018).
Подводя итог, можно сказать, что несколько исследований, изучающих острые эффекты физической активности как у грызунов, так и у мужчин, показали, что упражнения могут стимулировать как деградацию, так и синтез коллагена в скелетных мышцах.Восстановление микротравм, вызванных физической нагрузкой, следует двухфазному паттерну, при котором гликопротеины сначала создают переходную матрицу для управления катаболическими процессами, а анаболические процессы, укрепляющие структуру IMCT, происходят со значительной задержкой. Потенциал упражнений для ремоделирования ВКМ, по-видимому, зависит от режима сокращения, при этом эксцентрические сокращения вызывают большую реакцию, чем концентрическое или изометрическое мышечное действие. Доступно немного исследований, в которых проверяются результаты упражнений с различной интенсивностью, и пока результаты позволяют предположить, что распад белка (но не синтез) может быть сильнее спровоцирован более высокой интенсивностью.Неиспользование резко снижает активность ферментов, связанных с биосинтезом коллагенов, хотя на уровне белка изменения происходят с медленной скоростью. Поперечные сравнения с участием (в основном тренированных на выносливость) грызунов показывают, что хроническая физическая активность может привести к усилению фенотипа ММКИ. Единственные доступные на сегодняшний день долгосрочные продольные обучающие исследования были проведены на грызунах и предполагают, что длительные тренировки с отягощениями могут быть полезны для противодействия чрезмерному накоплению ММКИ в более старшем возрасте.Физиологические и функциональные последствия ремоделирования ММКИ, вызванного тренировкой, требуют дальнейшего изучения.
Заключение
Целью настоящего обзора является обзор современного состояния знаний о ECM скелетных мышц, который играет важную, хотя и часто недооцениваемую роль в поддержании мышечного гомеостаза, влияет на мышечную функцию и адаптацию и может быть ключом к лечению. мышечных и метаболических нарушений, вызванных старением или болезнью.
Как сложная сеть различных коллагенов, гликопротеинов, протеогликанов и эластина, ECM включает сократительные мышечные волокна и через интегрины и гликопротеиновый комплекс, связанный с дистрофином, служит биохимическим и механическим интерфейсом между мышечными клетками и их окружением. Сборке его коллагенового каркаса в основном способствуют факторы роста TGF-β и CTGF, которые регулируются различными протеогликанами, такими как декорин и бигликан. Более того, протеолитические ферменты (MMP), а также их ингибиторы (TIMP) участвуют в регуляции ECM.
Функционально ECM служит средой для передачи сократительной силы, которая может служить не только для повышения эффективности мышечного сокращения, но также для защиты мышечных волокон от чрезмерного напряжения и облегчения заживления микротравм. Помимо своей функциональной роли, ЕСМ активно участвует в регуляции пула сателлитных клеток в мышцах. Ниши ВКМ, образовавшиеся между сарколеммой и базальной мембраной, защищают сателлитные клетки от входа в клеточный цикл и, таким образом, помогают поддерживать регенеративный потенциал мышц.Конкретные компоненты ЕСМ, такие как фибронектин, коллаген VI и различные протеогликаны, могут дополнительно способствовать делению стволовых клеток. Напротив, ламинин, гликозаминогликаны и другие протеогликаны, как было показано, способствуют дифференцировке сателлитных клеток и их слиянию в зрелые миофибриллы.
Научные данные также демонстрируют, что ECM скелетных мышц является податливой тканью, которая может подвергаться процессам ремоделирования в результате старения, болезней, физических тренировок или неиспользования.В частности, старение обычно приводит к общему увеличению отложения коллагеновой ткани, изменениям в составе коллагена (переход к более высокому типу коллагена I к типу III) и увеличению неферментативного сшивания коллагена (за счет конечных продуктов гликирования). Эти изменения, которые, возможно, опосредованы снижением активности ММП, приводят к усилению жесткости ECM мышцы и могут ухудшить функцию и регенеративный потенциал мышцы.
Ремоделирование внеклеточного матрикса также может быть связано с метаболическими нарушениями, такими как диабет.Было обнаружено, что чрезмерное потребление пищи приводит к повышенной экспрессии генов, связанных с ECM (коллагены I, III, IV, V, SPARC, интегрин). В свою очередь, такое ремоделирование может нарушать передачу сигналов интегрина, тем самым снижая чувствительность к инсулину. Другими компонентами ECM, потенциально представляющими мишени для инсулинорезистентности, являются гиалуронан, комплекс дистрофин-дистрогликан, а также MMP9.
Наконец, ремоделирование ECM может быть вызвано физическими упражнениями. Хотя фактических исследований тренировок немного, есть доказательства того, что упражнения могут резко способствовать как увеличению синтеза коллагена (коллагены I, III, TGF-β1), так и деградации (MMP2, MMP9).Поперечные исследования на людях и продольные исследования на грызунах также предполагают, что такой повышенный обмен коллагена может привести к усилению коллагеновых структур у хронически тренированных субъектов и предотвратить чрезмерное отложение коллагена (то есть фиброз) в мышцах пожилого возраста. Исследования, изучающие последствия длительного неиспользования, показали противоречивые результаты. В то время как в ранних исследованиях сообщалось о снижении активности гидроксилазы и содержания гидроксипролина после кратковременной иммобилизации, в более поздних работах было обнаружено повышение содержания коллагена I после 21 дня разгрузки задних конечностей у крыс, но без изменений после 60 дней постельного режима у людей.Необходимы дальнейшие исследования и, в частности, обучение людей, чтобы изучить влияние различных методов обучения на структуру и состав ECM.
Авторские взносы
RC участвовал в литературных исследованиях и подготовил рукопись. MG и BW внесли свой вклад в литературные исследования и отредактировали рукопись. Все авторы одобрили окончательную версию рукописи и согласились нести ответственность за все аспекты работы. Все лица, обозначенные как авторы, имеют право на авторство, и все, кто имеет право на авторство, перечислены в списке.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы с благодарностью отмечаем финансовую поддержку этого исследования, полученную от Австрийского научного фонда (FWF): KLI 738-B27.
Список литературы
Aagaard, P., Andersen, J. L., Dyhre-Poulsen, P., Leffers, A.-M., Wagner, A., Магнуссон, С. П. и др. (2001). Механизм увеличения сократительной силы перистых мышц человека в ответ на силовую тренировку: изменения в архитектуре мышц. J. Physiol. 534, 613–623. DOI: 10.1111 / j.1469-7793.2001.t01-1-00613.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Адамс, Г. Р., Хазер, Б. М., Болдуин, К. М., и Дадли, Г. А. (1993). Состав тяжелых цепей миозина скелетных мышц и тренировки с отягощениями. J. Appl. Physiol. 74, 911–915. DOI: 10.1152 / jappl.1993.74.2.911
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Adstrum, S., Hedley, G., Schleip, R., Stecco, C., and Yucesoy, C.A. (2017). Определение фасциальной системы. J. Bodyw. Mov. Ther. 21, 173–177. DOI: 10.1016 / j.jbmt.2016.11.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Алмада, А. Э., и Вейджерс, А. Дж. (2016). Молекулярная схема судьбы стволовых клеток при регенерации скелетных мышц, старении и болезнях. Нат. Rev. Mol. Cell Biol. 17, 267–279. DOI: 10.1038 / nrm.2016.7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Американский колледж спортивной медицины. (2009). Позиционный стенд Американского колледжа спортивной медицины. Модели прогресса в тренировках с отягощениями для здоровых взрослых. Med. Sci. Спортивные упражнения. 41, 687–708. DOI: 10.1249 / mss.0b013e3181
0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Азизи, Э., Деслоре, А.Р., Холт, Н. К., Итон, К. Э. (2017). Устойчивость к радиальному расширению ограничивает мышечное напряжение и работу. Biomech. Модель. Механобиол. 16, 1633–1643. DOI: 10.1007 / s10237-017-0909-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Багдади, М. Б., Кастель, Д., Мачадо, Л., Фукада, С., Бирк, Д. Э., Релэкс, Ф. и др. (2018). Реципрокная передача сигналов с помощью Notch-Collagen V-CALCR удерживает мышечные стволовые клетки в их нише. Природа 557, 714–718. DOI: 10.1038 / s41586-018-0144-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Барретт Б. (1962). Длина и способ окончания отдельных мышечных волокон портняжной и задней бедренной мышцы человека. Acta Anat. 48, 242–257. DOI: 10.1159 / 000141843
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бенцингер, К. Ф., Ван, Ю. X., Дюмон, Н. А., и Рудницки, М. А. (2013a). Клеточная динамика в нише мышечных сателлитных клеток. EMBO Rep. 14, 1062–1072. DOI: 10.1038 / embor.2013.182
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бенцингер, К. Ф., Ван, Ю. Х., фон Мальцан, Дж., Сулеймани, В. Д., Инь, Х., и Рудницки, М. А. (2013b). Фибронектин регулирует передачу сигналов Wnt7a и рост сателлитных клеток. Стволовые клетки 12, 75–87. DOI: 10.1016 / j.stem.2012.09.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Берриа, Р., Ван, Л., Ричардсон, Д. К., Финлейсон, Дж., Белфорт, Р., Пратипанаватр, Т. и др. (2006). Повышенное содержание коллагена в инсулинорезистентных скелетных мышцах. Am. J. Physiol. Эндокринол. Метаб. 290, E560 – E565. DOI: 10.1152 / ajpendo.00202.2005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Блох, Р. Дж., И Гонсалес-Серратос, Х. (2003). Передача боковой силы через костамеры в скелетных мышцах. Exerc. Sport Sci. Ред. 31, 73–78. DOI: 10.1097 / 00003677-200304000-00004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Боонен, К.Дж. М., Розария-Чак, К. Ю., Баайенс, Ф. П. Т., ван дер Шафт, Д. В. Дж., И Пост, М. Дж. (2009). Важнейшие экологические сигналы из ниши сателлитных клеток: оптимизация пролиферации и дифференциации. Am. J. Physiol. Cell Physiol. 296, C1338 – C1345. DOI: 10.1152 / ajpcell.00015.2009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Боппарт, М. Д., и Махмассани, З. С. (2019). Передача сигналов интегрина: связь механической стимуляции с гипертрофией скелетных мышц. Am. J. Physiol. Cell Physiol. 317, C629 – C641. DOI: 10.1152 / ajpcell.00009.2019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брак А. С., Конбой И. М., Конбой М. Дж., Шен Дж. И Рандо Т. А. (2008). Временное переключение с передачи сигналов notch на Wnt в мышечных стволовых клетках необходимо для нормального миогенеза взрослых. Стволовые клетки клеток 2, 50–59. DOI: 10.1016 / j.stem.2007.10.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брак, А.С., Конбой, М. Дж., Рой, С., Ли, М., Куо, К. Дж., Келлер, К. и др. (2007). Повышенная передача сигналов Wnt во время старения изменяет судьбу мышечных стволовых клеток и увеличивает фиброз. Наука 317, 807–810. DOI: 10.1126 / science.1144090
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Brandan, E., Cabello-Verrugio, C., and Vial, C. (2008). Новые механизмы регуляции протеогликанов, декорина и бигликана во время формирования мышц и мышечной дистрофии. Matrix Biol. 27, 700–708. DOI: 10.1016 / j.matbio.2008.07.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брандт П. В., Лопес Э., Рубен Дж. П. и Грундфест Х. (1967). Взаимосвязь между плотностью упаковки миофиламентов и длиной саркомера в поперечно-полосатой мышце лягушки. J. Cell Biol. 33, 255–263. DOI: 10.1083 / jcb.33.2.255
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Браун С., Дэй С. и Доннелли А. (1999).Косвенные доказательства повреждения скелетных мышц человека и распада коллагена после эксцентрических действий мышц. J. Sports Sci. 17, 397–402. DOI: 10.1080 / 026404199365911
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бушби, К. М., Коллинз, Дж., И Хикс, Д. (2014). «Миопатии коллагенового типа VI», в Progress in Heritage Soft Connective Tissue Diseases , ed. Дж. Халпер, (Дордрехт: Спрингер), 185–199. DOI: 10.1007 / 978-94-007-7893-1_12
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Calve, S., Одельберг, С. Дж., И Саймон, Х.-Г. (2010). Переходный внеклеточный матрикс определяет поведение клеток во время регенерации мышц. Dev. Биол. 344, 259–271. DOI: 10.1016 / j.ydbio.2010.05.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кальво, А. К., Морено, Л., Морено, Л., Тойвонен, Дж. М., Манзано, Р., Молина, Н. и др. (2020). Коллаген типа XIX: перспективный биомаркер базальных мембран. Neural Regen. Res. 15, 988–995. DOI: 10.4103 / 1673-5374.270299
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кармели, Э., Моас, М., Леннон, С., и Пауэрс, С. К. (2005). Упражнения высокой интенсивности увеличивают экспрессию матричных металлопротеиназ в быстрых волокнах скелетных мышц: упражнения и ММП в быстрых волокнах скелета. Exp. Physiol. 90, 613–619. DOI: 10.1113 / expphysiol.2004.029462
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шарве, Б., Гиро, А., Malbouyres, M., Zwolanek, D., Guillon, E., Bretaud, S., et al. (2013). Нокдаун гена col22a1 у рыбок данио вызывает мышечную дистрофию за счет нарушения мышечно-сухожильного соединения. Dev. Camb. Англ. 140, 4602–4613. DOI: 10.1242 / dev.096024
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чике М., Бирк Д. Э., Беннеманн К. Г. и Кох М. (2014). Коллаген XII: защита целостности костей и мышц за счет организации фибрилл коллагена. Внутр. Дж.Биохим. Cell Biol. 53, 51–54. DOI: 10.1016 / j.biocel.2014.04.020
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кристенсен, С., Пурслоу, П. П. (2016). Роль матриксных металлопротеиназ в развитии мышечной и жировой ткани и качестве мяса: обзор. Meat Sci. 119, 138–146. DOI: 10.1016 / j.meatsci.2016.04.025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коркоран, М. Л., Хьюитт, Р. Э., Кляйнер, Д.Э. младший и Стетлер-Стивенсон У. Г. (1996). ММП-2: экспрессия, активация и ингибирование. Ферментный белок 49, 7–19. DOI: 10.1159 / 000468613
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Корнелисон Д. Д., Филла М. С., Стэнли Х. М., Рапрэгер А. С. и Олвин Б. Б. (2001). Синдекан-3 и синдекан-4 специфически маркируют сателлитные клетки скелетных мышц и участвуют в поддержании сателлитных клеток и регенерации мышц. Dev. Биол. 239, 79–94.DOI: 10.1006 / dbio.2001.0416
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Косгроув, Б. Д., Гилберт, П. М., Порпилья, Э., Муркиоти, Ф., Ли, С. П., Корбел, С. Ю. и др. (2014). Омоложение популяции мышечных стволовых клеток восстанавливает силу травмированных старых мышц. Нат. Med. 20, 255–264. DOI: 10,1038 / нм 3464
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Crameri, R.M., Aagaard, P., Qvortrup, K., Langberg, H., Олесен, Дж., И Кьяер, М. (2007). Повреждение миофибрилл в скелетных мышцах человека: эффекты электростимуляции против произвольных сокращений: равномерная против гетерогенная активация мышечных волокон. J. Physiol. 583, 365–380. DOI: 10.1113 / jphysiol.2007.128827
CrossRef Полный текст | Google Scholar
Крамери, Р. М., Лангберг, Х., Тейснер, Б., Магнуссон, П., Шредер, Х. Д., Олесен, Дж. Л. и др. (2004). Усиление обработки проколлагена в скелетных мышцах после однократной эксцентрической нагрузки у людей. Matrix Biol. 23, 259–264. DOI: 10.1016 / j.matbio.2004.05.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цуй Н., Ху М. и Халил Р. А. (2017). Биохимические и биологические признаки матричных металлопротеиназ. Прог. Мол. Биол. Пер. Sci. 147, 1–73. DOI: 10.1016 / bs.pmbts.2017.02.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
де Соуза Нето, И. В., Дуриган, Дж. Л. К., Гуццони, В., Тибана, Р.A., Prestes, J., de Araujo, H. S. S. и др. (2018). Влияние тренировок с отягощениями на активность матричной металлопротеиназы в скелетных мышцах и кровообращение при старении. Фронт. Physiol. 9: 190. DOI: 10.3389 / fphys.2018.00190
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дитрих А. В., Боттер А., Виейра Т. М., Пеолссон А., Петцке Ф., Дэйви П. и др. (2017). Пространственная вариация и несоответствие между оценками начала мышечной активации по ЭМГ и УЗИ. Sci. Отчет 7: 42011. DOI: 10.1038 / srep42011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Dietrich, S., Abou-Rebyeh, F., Brohmann, H., Bladt, F., Sonnenberg-Riethmacher, E., Yamaai, T., et al. (1999). Роль SF / HGF и c-Met в развитии скелетных мышц. Dev. Camb. Англ. 126, 1621–1629.
PubMed Аннотация | Google Scholar
Duance, V.C., Restall, D.J., Beard, H., Bourne, F.J, and Bailey, A.J. (1977). Расположение трех типов коллагена в скелетных мышцах. FEBS Lett. 79, 248–252. DOI: 10.1016 / 0014-5793 (77) 80797-7
CrossRef Полный текст | Google Scholar
Эклунд, Л., Пиухола, Дж., Комулайнен, Дж., Сормунен, Р., Онгваррасопоне, К., Фасслер, Р. и др. (2001). Недостаток коллагена типа XV вызывает у мышей скелетную миопатию и сердечно-сосудистые дефекты. Proc. Natl. Акад. Sci. США 98, 1194–1199. DOI: 10.1073 / pnas.031444798
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Энглер, А.Дж., Гриффин, М. А., Сен, С., Беннеман, К. Г., Суини, Х. Л. и Дишер, Д. Е. (2004). Миотрубки оптимально дифференцируются на субстратах с тканевой жесткостью: патологические последствия для мягких или жестких микросред. J. Cell Biol. 166, 877–887. DOI: 10.1083 / jcb.200405004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Этьен, Дж., Лю, К., Скиннер, К. М., Конбой, М. Дж., И Конбой, И. М. (2020). Скелетные мышцы как экспериментальная модель выбора для изучения старения и омоложения тканей. Скелет. Мышца 10: 4. DOI: 10.1186 / s13395-020-0222-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фишер, Дж., Стил, Дж., Брюс-Лоу, С., и Смит, Д. (2011). Рекомендации по тренировкам с отягощениями, основанные на фактах. Med. Спорт. 15, 147–162. DOI: 10.2478 / v10036-011-0025-x
CrossRef Полный текст | Google Scholar
Фолланд, Дж. П., и Уильямс, А. Г. (2007). Адаптация к силовым тренировкам: морфологический и неврологический вклад в увеличение силы. Sports Med. 37, 145–168. DOI: 10.2165 / 00007256-200737020-00004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фрай, К. С., Кирби, Т. Дж., Космак, К., Маккарти, Дж. Дж., И Петерсон, К. А. (2017). Миогенные клетки-предшественники контролируют продукцию внеклеточного матрикса фибробластами во время гипертрофии скелетных мышц. Стволовые клетки клеток 20, 56–69. DOI: 10.1016 / j.stem.2016.09.010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гарбер, К.E., Blissmer, B., Deschenes, M. R., Franklin, B. A., Lamonte, M. J., Lee, I.-M., et al. (2011). Количество и качество упражнений для развития и поддержания кардиореспираторной, скелетно-мышечной и нейромоторной формы у практически здоровых взрослых: руководство по назначению упражнений. Med. Sci. Спортивные упражнения. 43, 1334–1359. DOI: 10.1249 / MSS.0b013e318213fefb
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гилберт, П. М., Хэвенстрайт, К. Л., Магнуссон, К.Э. Г., Сакко А., Леонарди Н. А., Крафт П. и др. (2010). Эластичность субстрата регулирует самообновление стволовых клеток скелетных мышц в культуре. Science 329, 1078–1081. DOI: 10.1126 / science.11
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гиллис, А. Р., Чепмен, М. А., Бушонг, Э. А., Деринк, Т. Дж., Эллисман, М. Х., и Либер, Р. Л. (2017). Трехмерная реконструкция с высоким разрешением внеклеточного матрикса фиброзных скелетных мышц: организация внеклеточного матрикса фиброзных мышц. J. Physiol. 595, 1159–1171. DOI: 10.1113 / JP273376
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Госселин, Л. Е., Адамс, К., Коттер, Т. А., Маккормик, Р. Дж., И Томас, Д. П. (1998). Влияние физической тренировки на пассивной жесткости в опорно-двигательном аппарате скелетных мышц: роль внеклеточного матрикса. J. Appl. Physiol. 85, 1011–1016. DOI: 10.1152 / jappl.1998.85.3.1011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Грефте, С., Vullinghs, S., Kuijpers-Jagtman, A.M., Torensma, R., and Von den Hoff, J. W. (2012). Матригель, но не коллаген I, поддерживает способность к дифференцировке мышечных клеток in vitro . Biomed. Матер. 7: 055004. DOI: 10.1088 / 1748-6041 / 7/5/055004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Grounds, M. D. (1998). Возрастные изменения реакции клеток скелетных мышц на упражнения и регенерацию. Ann. Акад.Sci. 854, 78–91. DOI: 10.1111 / j.1749-6632.1998.tb09894.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Основание, М. Д., Сорокин, Л., Уайт, Дж. (2005). Сила на границе межклеточного матрикса и мышцы. Сканд. J. Med. Sci. Спорт 15, 381–391. DOI: 10.1111 / j.1600-0838.2005.00467.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гийон, Э., Брето, С., Руджеро, Ф. (2016). Предшественники медленных мышц откладывают отпечаток пальца матрицы XV коллагена, чтобы направлять навигацию по моторным аксонам. J. Neurosci. 36, 2663–2676. DOI: 10.1523 / JNEUROSCI.2847-15.2016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гутьеррес, Дж., И Брандан, Э. (2010). Новый механизм связывания фактора роста фибробластов 2 с помощью глипикана в липидных рафтах, позволяющий дифференцировать скелетные мышцы. Мол. Клетка. Биол. 30, 1634–1649. DOI: 10.1128 / mcb.01164-09
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гуццони, В., Рибейро, М.Б. Т., Лопес, Г. Н., де Касия Маркети, Р., де Андраде, Р. В., Селистр-де-Араужо, Х. С. и др. (2018). Влияние тренировки с отягощениями на адаптацию внеклеточного матрикса в скелетных мышцах старых крыс. Фронт. Physiol. 9: 374. DOI: 10.3389 / fphys.2018.00374
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Халпер, Дж., И Кьяер, М. (2014). «Основные компоненты соединительной ткани и внеклеточного матрикса: эластин, фибриллин, фибулины, фибриноген, фибронектин, ламинин, тенасцины и тромбоспондины», в Progress in Heritable Soft Connective Tissue Diseases , ed.Дж. Халпер, (Дордрехт: Спрингер), 31–47. DOI: 10.1007 / 978-94-007-7893-1_3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Härönen, H., Zainul, Z., Tu, H., Naumenko, N., Sormunen, R., Miinalainen, I., et al. (2017). Коллаген XIII обеспечивает пре- и постсинаптическую целостность нервно-мышечного синапса. Hum. Мол. Genet. 26, 2076–2090. DOI: 10.1093 / hmg / ddx101
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хассон П., ДеЛорье А., Беннетт М., Григорьева Э., Найче Л. А., Папайоанну В. Э. и др. (2010). Tbx4 и tbx5, действующие в соединительной ткани, необходимы для формирования рисунка мышц конечностей и сухожилий. Dev. Cell 18, 148–156. DOI: 10.1016 / j.devcel.2009.11.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хаус, Дж. М., Карритерс, Дж. А., Кэрролл, К. К., Теш, П. А., и Трапп, Т. А. (2007a). Содержание белка сократительной и соединительной ткани в скелетных мышцах человека: эффекты 35- и 90-дневного моделирования микрогравитации и контрмеры при физической нагрузке. Am. J. Physiol. Regul. Интегр. Комп. Physiol. 293, R1722 – R1727. DOI: 10.1152 / ajpregu.00292.2007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хаус, Дж. М., Карритерс, Дж. А., Траппе, С. У. и Траппе, Т. А. (2007b). Коллаген, сшивание и конечные продукты гликирования в стареющих скелетных мышцах человека. J. Appl. Physiol. 103, 2068–2076. DOI: 10.1152 / japplphysiol.00670.2007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хейккинен, А., Härönen, H., Norman, O., and Pihlajaniemi, T. (2019). Коллаген XIII и другие компоненты ECM в сборке и заболевании нервно-мышечного соединения. Анат. Рек. doi: 10.1002 / ar.24092 [Epub перед печатью].
CrossRef Полный текст | PubMed Аннотация | Google Scholar
Heinemeier, K. M., Olesen, J. L., Haddad, F., Langberg, H., Kjaer, M., Baldwin, K. M., et al. (2007). Экспрессия коллагена и родственных факторов роста в сухожилиях и скелетных мышцах крысы в ответ на определенные типы сокращения: экспрессия коллагена и TGF-β-1 в тренированных сухожилиях и мышцах. J. Physiol. 582, 1303–1316. DOI: 10.1113 / jphysiol.2007.127639
CrossRef Полный текст | Google Scholar
Heljasvaara, R., Aikio, M., Ruotsalainen, H., and Pihlajaniemi, T. (2017). Коллаген XVIII в гомеостазе и нарушении регуляции тканей — уроки, извлеченные из модельных организмов и пациентов-людей. Matrix Biol. 57–58, 55–75. DOI: 10.1016 / j.matbio.2016.10.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хидзиката, Т., Вакисака Х. и Ниида С. (1993). Функциональная комбинация сужающихся профилей и наложений в нерасширяющихся волокнах скелетных мышц, оканчивающихся внутрипучко. Анат. Рек. 236, 602–610. DOI: 10.1002 / ar.10403
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хиндл, А.Г., Хорнинг, М., Меллиш, Дж .-А. Э. и Лоулер Дж. М. (2009). Погружение в старость: мускульное старение у крупнотелого долгоживущего млекопитающего, тюленя Уэдделла ( Leptonychotes weddellii ). J. Exp. Биол. 212, 790–796. DOI: 10.1242 / jeb.025387
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Холм, Л., Рахбек, С. К., Фаруп, Дж., Вендельбо, М. Х., и Виссинг, К. (2017). Режим сокращения и потребление сывороточного протеина влияют на скорость синтеза внутримышечной соединительной ткани: краткие отчеты. Мышечный нерв 55, 128–130. DOI: 10.1002 / mus.25398
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Холм, Л., ван Холл, Г., Роуз, А. Дж., Миллер, Б. Ф., Дёссинг, С., Рихтер, Э. А. и др. (2010). Интенсивность сокращения и питание по-разному влияют на скорость синтеза коллагена и миофибриллярного белка в скелетных мышцах человека. Am. J. Physiol. Эндокринол. Метаб. 298, E257 – E269. DOI: 10.1152 / ajpendo.00609.2009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хуанг, Ю., Фан, Ю., Саланова, М., Ян, X., Сан, Л., и Блоттнер, Д. (2018). Влияние подошвенной вибрации на кости и глубокую фасцию в модели неиспользования разгрузки задней конечности крысы. Фронт. Physiol. 9: 616. DOI: 10.3389 / fphys.2018.00616
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хьюз, Д. К., Маркотт, Г. Р., Маршалл, А. Г., Уэст, Д. В. Д., Баер, Л. М., Уоллес, М. А. и др. (2016). Возрастные различия дистрофина: влияние на белки передачи силы, целостность мембран и стабильность нервно-мышечных соединений. J. Gerontol. A. Biol. Sci. Med. Sci. 72, 640–648. DOI: 10.1093 / gerona / glw109
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хуцзин, П.А. (1999). Мышцы как композит, армированный коллагеновыми волокнами: обзор передачи силы в мышце и всей конечности. J. Biomech. 32, 329–345. DOI: 10.1016 / s0021-9290 (98) 00186-9
CrossRef Полный текст | Google Scholar
Хьюцзин, П. А., Баан, Г. К., и Ребел, Г. Т. (1998). Немышухожильная передача силы в длинном разгибателе пальцев большого пальца крысы. J. Exp. Биол. 201, 683–691.
Google Scholar
Хилдал, Р. Д., Нельсон, Б., Xin, L., Welling, T., Groscost, L., Hubal, M.J., et al. (2015). Ремоделирование внеклеточного матрикса и его вклад в защитную адаптацию после удлинения сокращений в мышцах человека. FASEB J. 29, 2894–2904. DOI: 10.1096 / fj.14-266668
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ивата, Дж., Судзуки, А., Пеликан, Р. К., Хо, Т.-В., и Чай, Ю. (2013). Передача сигналов неканонического трансформирующего фактора роста β (TGFβ) в клетках краниального нервного гребня вызывает дефекты развития мышц языка. J. Biol. Chem. 288, 29760–29770. DOI: 10.1074 / jbc.M113.493551
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Якобсен, Дж. Р., Макки, А. Л., Кнудсен, А. Б., Кох, М., Кьяер, М., и Крогсгаард, М. Р. (2017). Состав и адаптация мышечно-сухожильных суставов человека и соседних мышечных волокон к тяжелым тренировкам с отягощениями. Сканд. J. Med. Sci. Спорт 27, 1547–1559. DOI: 10.1111 / sms.12794
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Яначек, Я., Цебашек, В., Кубинова, Л., Рибарич, С., и Эрзен, И. (2009). Трехмерная визуализация и измерение капилляров, снабжающих метаболически различные типы волокон в длинном разгибателе пальцев большого пальца крысы во время денервации и реиннервации. J. Histochem. Cytochem. 57, 437–447. DOI: 10.1369 / jhc.2008.953018
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ярвинен, Т. А. Х., Йожа, Л., Каннус, П., Ярвинен, Т. Л. Н., и Ярвинен, М. (2002). Организация и распределение внутримышечной соединительной ткани в нормальных и неподвижных скелетных мышцах.Иммуногистохимическое, поляризационное и сканирующее электронно-микроскопические исследования. J. Muscle Res. Cell Motil. 23, 245–254. DOI: 10.1023 / A: 1020
8336
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Канг, Л., Лантье, Л., Кеннеди, А., Боннер, Дж. С., Мэйс, В. Х., Брейси, Д. П. и др. (2013). Гиалуронан накапливается при кормлении с высоким содержанием жиров и способствует развитию инсулинорезистентности. Диабет 62, 1888–1896. DOI: 10.2337 / db12-1502
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Канг, Л., Мэйс, У. Х., Джеймс, Ф. Д., Брейси, Д. П., и Вассерман, Д. Х. (2014). Матричная металлопротеиназа 9 противодействует индуцированной диетой мышечной инсулинорезистентности у мышей. Diabetologia 57, 603–613. DOI: 10.1007 / s00125-013-3128-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кардон Г., Харф Б. Д. и Табин К. Дж. (2003). Tcf4-позитивная мезодермальная популяция обеспечивает предпаттерн для формирования мышечного паттерна конечностей позвоночных. Dev. Cell 5, 937–944. DOI: 10.1016 / с 1534-5807 (03) 00360-5
CrossRef Полный текст | Google Scholar
Карпакка, Й., Вяэнянен, К., Орава, С., и Такала, Т. Э. (1990). Эффекты предварительной иммобилизации и иммобилизации на синтез коллагена в скелетных мышцах крыс. Внутр. J. Sports Med. 11, 484–488. DOI: 10.1055 / с-2007-1024842
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Карпакка, Дж., Виртанен, П., Ваананен, К., Орава, С., и Такала, Т. Е. (1991).Синтез коллагена в скелетных мышцах крысы во время иммобилизации и ремобилизации. J. Appl. Physiol. 70, 1775–1780. DOI: 10.1152 / jappl.1991.70.4.1775
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Khaleduzzaman, M., Sumiyoshi, H., Ueki, Y., Inoguchi, K., Ninomiya, Y., and Yoshioka, H. (1997). Структура гена коллагена человека типа XIX (COL19A1), которая предполагает, что он произошел от гена-предка семейства FACIT. Геномика 45, 304–312.DOI: 10.1006 / geno.1997.4921
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ким Дж., Ван З., Хеймсфилд С. Б., Баумгартнер Р. Н. и Галлахер Д. (2002). Общая масса скелетных мышц: оценка с помощью нового метода двухэнергетической рентгеновской абсорбциометрии. Am. J. Clin. Nutr. 76, 378–383. DOI: 10.1093 / ajcn / 76.2.378
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кнудсен, А.Б., Ларсен, М., Макки, А.Л., Хьорт, М., Hansen, K. K., Qvortrup, K., et al. (2015). Человеческое мышечно-сухожильное соединение: ультраструктурное исследование и трехмерный анализ. Сканд. J. Med. Sci. Sports 25, e116 – e123. DOI: 10.1111 / sms.12221
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Koch, M., Schulze, J., Hansen, U., Ashwodt, T., Keene, D.R., Brunken, W.J., et al. (2004). Новый маркер тканевых соединений, коллаген XXII. J. Biol. Chem. 279, 22514–22521. DOI: 10.1074 / jbc.M400536200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кованен, В., Суоминен, Х., Ристели, Дж., И Ристели, Л. (1988). Коллаген IV типа и ламинин в медленных и быстрых скелетных мышцах крыс — Влияние возраста и тренировок на выносливость на протяжении всей жизни. Coll. Relat. Res. 8, 145–153. DOI: 10,1016 / s0174-173x (88) 80026-8
CrossRef Полный текст | Google Scholar
Куо, Х. Дж., Маслен, К. Л., Кин, Д. Р., и Гланвилл, Р. У. (1997). Коллаген VI типа закрепляет базальные мембраны эндотелия, взаимодействуя с коллагеном IV типа. J. Biol. Chem. 272, 26522–26529. DOI: 10.1074 / jbc.272.42.26522
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Леонг, Д. П., Тео, К. К., Рангараджан, С., Лопес-Харамильо, П., Авесум, А., Орландини, А., и др. (2015). Прогностическое значение силы хвата: результаты проспективного исследования городской и сельской эпидемиологии (PURE). Lancet Lond. Англ. 386, 266–273. DOI: 10,1016 / s0140-6736 (14) 62000-6
CrossRef Полный текст | Google Scholar
Ли, Дж.Дж., Виттерт, Г. А., Винсент, А., Атлантис, Э., Ши, З., Эпплтон, С. Л. и др. (2016). Сила мышечного захвата позволяет прогнозировать развитие диабета 2 типа: популяционное когортное исследование. Метаболизм 65, 883–892. DOI: 10.1016 / j.metabol.2016.03.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Либер, Р. Л., и Уорд, С. Р. (2013). Клеточные механизмы тканевого фиброза. 4. Структурные и функциональные последствия фиброза скелетных мышц. Am. J. Physiol.Cell Physiol. 305, C241 – C252. DOI: 10.1152 / ajpcell.00173.2013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лопес-Отин, К., Бласко, М. А., Партридж, Л., Серрано, М., и Кремер, Г. (2013). Признаки старения. Cell 153, 1194–1217. DOI: 10.1016 / j.cell.2013.05.039
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Maaß, T., Bayley, C.P., Mörgelin, M., Lettmann, S., Bonaldo, P., Paulsson, M., et al. (2016).Неоднородность микрофибрилл коллагена VI: структурный анализ неколлагеновых участков. J. Biol. Chem. 291, 5247–5258. DOI: 10.1074 / jbc.M115.705160
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Mackey, A. L., Brandstetter, S., Schjerling, P., Bojsen-Moller, J., Qvortrup, K., Pedersen, M. M., et al. (2011). Последовательный ответ мертвых клеток внеклеточного матрикса и фиброзных регуляторов после повреждения мышц участвует в защите скелетных мышц человека от будущих повреждений. FASEB J. 25, 1943–1959. DOI: 10.1096 / fj.10-176487
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Mackey, A. L., Donnelly, A. E., Turpeenniemi-Hujanen, T., and Roper, H.P. (2004). Содержание коллагена в скелетных мышцах у людей после эксцентрических сокращений большой силы. J. Appl. Physiol. 97, 197–203. DOI: 10.1152 / japplphysiol.01174.2003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Махновский П.А., Згода В.Г., Боков Р.О., Шагимарданова Е.И., Газизова Г.Р., Гусев О.А. и др. (2020). Регуляция белков в скелетных мышцах человека: роль транскрипции. Sci. Отчет 10: 3514.
Google Scholar
Мартин, Г. Р., и Тимпл, Р. (1987). Ламинин и другие компоненты базальной мембраны. Annu. Rev. Cell Biol. 3, 57–85. DOI: 10.1146 / annurev.cb.03.110187.000421
CrossRef Полный текст | Google Scholar
Макки, Т.Дж., Перлман, Г., Моррис, М., Комарова, С. В. (2019). Состав внеклеточного матрикса соединительной ткани: систематический обзор и метаанализ. Sci. Отчет 9: 10542.
Google Scholar
Мендиас, К. Л., Шварц, А. Дж., Грекин, Дж. А., Гумусио, Дж. П., и Сугг, К. Б. (2017). Изменения сократимости мышечных волокон и продукции внеклеточного матрикса при гипертрофии скелетных мышц. J. Appl. Physiol. 122, 571–579. DOI: 10.1152 / japplphysiol.00719.2016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Монти, Р.Дж., Рой, Р. Р., Ходжсон, Дж. А., и Реджи Эджертон, В. (1999). Передача сил в скелетных мышцах млекопитающих. J. Biomech. 32, 371–380. DOI: 10.1016 / s0021-9290 (98) 00189-4
CrossRef Полный текст | Google Scholar
Мур, Д. Р., Филлипс, С. М., Бабрадж, Дж. А., Смит, К., и Ренни, М. Дж. (2005). Синтез миофибриллярных и коллагеновых белков в скелетных мышцах человека у молодых мужчин после максимального сокращения и удлинения сокращений. Am. J. Physiol.Эндокринол. Метаб. 288, E1153 – E1159. DOI: 10.1152 / ajpendo.00387.2004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Моралес М.Г., Кабелло-Верругио К., Сантандер К., Кабрера Д., Гольдшмединг Р. и Брандан Э. (2011). Сверхэкспрессия CTGF / CCN-2 может напрямую вызывать признаки дистрофии скелетных мышц. J. Pathol. 225, 490–501. DOI: 10.1002 / путь.2952
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Малви, К., Харно, Э., Кинан, А., и Олендик, К. (2005). Экспрессия комплекса дистрофин-дистрогликан скелетных мышц и комплекса синтрофин-оксид азота серьезно нарушена у крыс Goto-Kakizaki с диабетом 2 типа. Eur. J. Cell Biol. 84, 867–883. DOI: 10.1016 / j.ejcb.2005.06.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Наба А., Клаузер К. Р., Динг Х., Уиттакер К. А., Карр С. А. и Хайнс Р. О. (2016). Внеклеточный матрикс: инструменты и идеи для эпохи «омиков». Matrix Biol. 49, 10–24. DOI: 10.1016 / j.matbio.2015.06.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нассари, С., Дюпре, Д., Фурнье-Тибо, К. (2017). Немиогенный вклад в развитие мышц и гомеостаз: роль соединительных тканей. Фронт. Cell Dev. Биол. 5:22. DOI: 10.3389 / fcell.2017.00022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Nordez, A., Gallot, T., Catheline, S., Гевель А., Корню К. и Хуг Ф. (2009). Повторно рассмотрена электромеханическая задержка с использованием ультразвука с очень высокой частотой кадров. J. Appl. Physiol. 106, 1970–1975. DOI: 10.1152 / japplphysiol.00221.2009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Новицки Дж. Л., Такимото Р. и Берк А. С. (2003). Латеральная сомитная граница: дорсо-вентральные аспекты передне-задней регионализации у эмбрионов птиц. мех. Dev. 120, 227–240. DOI: 10.1016 / s0925-4773 (02) 00415-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Огасавара, Р., Наказато К., Сато К., Боппарт М. Д. и Фудзита С. (2014). Упражнения с отягощениями увеличивают экспрессию активного белка MMP и бета1-интегрина в скелетных мышцах. Physiol. Отчет 2: e12212. DOI: 10.14814 / phy2.12212
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Olguin, H.C., и Olwin, B.B. (2004). Повышающая регуляция Pax-7 ингибирует миогенез и прогрессию клеточного цикла в сателлитных клетках: потенциальный механизм самообновления. Dev. Биол. 275, 375–388.DOI: 10.1016 / j.ydbio.2004.08.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Олссон, Л., Фальк, П., Лопес, К., Кобб, Дж., И Ханкен, Дж. (2001). Клетки черепного нервного гребня вносят вклад в соединительную ткань в черепных мышцах бесхвостых амфибий, Bombina orientalis . Dev. Биол. 237, 354–367. DOI: 10.1006 / dbio.2001.0377
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Passerieux, E., Rossignol, R., Летелье, Т., и Делаж, Дж. (2007). Физическая непрерывность перимизиума от миофибрилл к сухожилиям: участие в передаче латеральной силы в скелетных мышцах. J. Struct. Биол. 159, 19–28. DOI: 10.1016 / j.jsb.2007.01.022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Паттисон, Дж. С., Фолк, Л. К., Мэдсен, Р. У., Чайлдс, Т. Э. и Бут, Ф. У. (2003). Транскрипционное профилирование выявляет обширное подавление экспрессии гена внеклеточного матрикса в саркопенической камбаловидной мышце крысы. Physiol. Геномика 15, 34–43. DOI: 10.1152 / Physiolgenomics.00040.2003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Паван, П. Г., Стекко, А., Стерн, Р., и Стекко, К. (2014). Болезненные связи: уплотнение или фиброз фасции. Curr. Боль Головная боль Реп. 18: 441.
Google Scholar
Пирс Р. В., Шерц П. Дж., Кэмпбелл Дж. К. и Табин К. Дж. (2007). Анализ клеточного происхождения зачатка куриной конечности. Dev. Биол. 310, 388–400. DOI: 10.1016 / j.ydbio.2007.08.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Питер А. К., Ченг Х., Росс Р. С., Ноултон К. У. и Чен Дж. (2011). Костамер соединяет саркомеры с сарколеммой поперечно-полосатой мышцы. Прог. Педиатр. Кардиол. 31, 83–88. DOI: 10.1016 / j.ppedcard.2011.02.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пурслоу П. П. (2002). Структура и функциональное значение изменений соединительной ткани в мышцах. Комп. Биохим. Physiol. Мол. Интегр. Physiol. 133, 947–966. DOI: 10.1016 / s1095-6433 (02) 00141-1
CrossRef Полный текст | Google Scholar
Рамасвами, К. С., Палмер, М. Л., ван дер Мейлен, Дж. Х., Рену, А., Костроминова, Т. Ю., Мишель, Д. Е. и др. (2011). Боковая передача силы нарушена в скелетных мышцах дистрофических мышей и очень старых крыс: боковая передача силы в скелетных мышцах мышей и крыс. J. Physiol. 589, 1195–1208.DOI: 10.1113 / jphysiol.2010.201921
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Райагири, С.С., Ранальди, Д., Рэйвен, А., Мохамад Азхар, Н.И.Ф., Лефевр, О., Заммит, П.С., и др. (2018). Ремоделирование базальной пластинки в нише стволовых клеток скелетных мышц опосредует самообновление стволовых клеток. Нат. Commun. 9: 1075.
Google Scholar
Риз, С. П., Андервуд, К. Дж., И Вайс, Дж. А. (2013). Влияние протеогликана декорина на фибриллогенез, ультраструктуру и механику коллагеновых гелей типа I. Matrix Biol. 32, 414–423. DOI: 10.1016 / j.matbio.2013.04.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Реннинг, С. Б., Педерсен, М. Э., Андерсен, П. В., и Холлунг, К. (2013). Комбинация гликозаминогликанов и фиброзных белков улучшает пролиферацию клеток и раннюю дифференциацию первичных клеток скелетных мышц крупного рогатого скота. Дифференциация 86, 13–22. DOI: 10.1016 / j.diff.2013.06.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роуленд, Л.А., Бал Н. С., Периасами М. (2015). Роль термогенных механизмов скелетных мышц в эндотермии позвоночных. Biol. Преподобный Камб. Филос. Soc. 90, 1279–1297. DOI: 10.1111 / brv.12157
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рубенштейн, А. Б., Смит, Г. Р., Рауэ, У., Бег, Г., Минчев, К., Руф-Замойски, Ф. и др. (2020). Одноклеточные профили транскрипции в скелетных мышцах человека. Sci. Отчет 10: 229.
Google Scholar
Сабателли, П., Гуаланди, Ф., Гара, С. К., Грумати, П., Зампарелли, А., Мартони, Э. и др. (2012). Экспрессия α5 и α6 цепей коллагена VI в мышцах человека и при мышечном фиброзе, связанном с мышечной дистрофией Дюшенна. Matrix Biol. 31, 187–196. DOI: 10.1016 / j.matbio.2011.12.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Санес, Дж. Р. (1982). Ламинин, фибронектин и коллаген в синаптических и внесинаптических частях базальной мембраны мышечных волокон. Дж.Cell Biol. 93, 442–451. DOI: 10.1083 / jcb.93.2.442
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Саволайнен, Дж., Вяэнянен, К., Пуранен, Дж., Такала, Т. Э., Комулайнен, Дж., И Вихко, В. (1988). Синтез коллагена и протеолитическая активность в скелетных мышцах крыс: эффект гипсовой иммобилизации в удлиненных и укороченных положениях. Arch. Phys. Med. Rehabil. 69, 964–969.
PubMed Аннотация | Google Scholar
Шенфельд, Б.Дж. (2010). Механизмы гипертрофии мышц и их применение в тренировках с отягощениями. J. Strength Cond. Res. 24, 2857–2872. DOI: 10.1519 / JSC.0b013e3181e840f3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Schoenrock, B., Zander, V., Dern, S., Limper, U., Mulder, E., Veraksitš, A., et al. (2018). Постельный режим, противодействие физическим упражнениям и восстанавливающее воздействие на систему мышечного тонуса человека в состоянии покоя. Фронт. Physiol. 9: 810. DOI: 10.3389 / fphys.2018.00810
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Schönherr, E., Witsch-Prehm, P., Harrach, B., Robenek, H., Rauterberg, J., and Kresse, H. (1995). Взаимодействие бигликана с коллагеном I типа. J. Biol. Chem. 270, 2776–2783. DOI: 10.1074 / jbc.270.6.2776
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шварц Р. И. (2015). Коллаген I и фибробласты: высокая экспрессия белка требует новой парадигмы посттранскрипционной регуляции с обратной связью. Biochem. Биофиз. Rep. 3, 38–44. DOI: 10.1016 / j.bbrep.2015.07.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сефтон, Э. М., и Кардон, Г. (2019). Связующее развитие мышц, врожденные дефекты и эволюция: важная роль соединительной ткани мышц. Curr. Верхний. Dev. Биол. 132, 137–176. DOI: 10.1016 / bs.ctdb.2018.12.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шарафи Б. и Блемкер С.С. (2011). Математическая модель передачи силы от оканчивающихся внутри ячеек мышечных волокон. J. Biomech. 44, 2031–2039. DOI: 10.1016 / j.jbiomech.2011.04.038
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Соренсен, Дж. Р., Скоузен, К., Холланд, А., Уильямс, К., и Хилдал, Р. Д. (2018). Острый внеклеточный матрикс, воспалительная реакция и реакция MAPK на удлинение сокращений в скелетных мышцах пожилого человека. Exp. Геронтол. 106, 28–38.DOI: 10.1016 / j.exger.2018.02.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Street, S. F. (1983). Боковая передача напряжения в миофибриллах лягушки: миофибриллярная сеть и поперечные цитоскелетные связи являются возможными передатчиками. J. Cell. Physiol. 114, 346–364. DOI: 10.1002 / jcp.1041140314
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сумиёси, Х., Лауб, Ф., Йошиока, Х., и Рамирес, Ф. (2001).Эмбриональная экспрессия коллагена типа XIX преходяща и ограничена мышечными клетками. Dev. Дин. 220, 155–162. DOI: 10.1002 / 1097-0177 (2000) 9999: 9999 <:: aid-dvdy1099> 3.0.co; 2-w
CrossRef Полный текст | Google Scholar
Суоминен, Х., Хейккинен, Э. (1975). Активность ферментов в мышечной и соединительной ткани M. Vastus lateralis у мужчин в возрасте от 33 до 70 лет, которые обычно тренируются и ведут малоподвижный образ жизни. Eur. J. Appl. Physiol. 34, 249–254. DOI: 10.1007 / bf00999938
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Суоминен, Х., Хейккинен, Э. и Паркатти, Т. (1977). Влияние восьминедельной физической подготовки на мышцы и соединительную ткань M. Vastus lateralis у 69-летних мужчин и женщин. J. Gerontol. 32, 33–37. DOI: 10.1093 / geronj / 32.1.33
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Swartz, M. E., Eberhart, J., Pasquale, E. B., and Krull, C.Э. (2001). Взаимодействия EphA4 / эфрин-A5 при миграции клеток-предшественников мышц в передних конечностях птиц. Dev. Camb. Англ. 128, 4669–4680.
PubMed Аннотация | Google Scholar
Такаги Р., Огасавара Р., Цутаки А., Наказато К. и Исии Н. (2016). Региональная адаптация коллагена в скелетных мышцах к повторяющимся сериям напряженных эксцентрических упражнений. Pflugers Arch. 468, 1565–1572. DOI: 10.1007 / s00424-016-1860-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Такала, Т.Э. и Виртанен П. (2000). Биохимический состав внеклеточного матрикса мышц: эффект нагрузки. Сканд. J. Med. Sci. Спорт 10, 321–325. DOI: 10.1034 / j.1600-0838.2000.010006321.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Такала Т. Е., Мюллюля Р., Салминен А., Анттинен Х. и Вихко В. (1983). Повышенная активность пролил-4-гидроксилазы и галактозилгидроксилизилглюкозилтрансферазы, ферментов биосинтеза коллагена, в скелетных мышцах мышей, тренированных на выносливость. Pflügers Arch. 399, 271–274. DOI: 10.1007 / BF00652751
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Там, К. С., Чаудхури, Р., Хатчисон, А. Т., Самоча-Бонет, Д., и Хайльбронн, Л. К. (2017). Ремоделирование внеклеточного матрикса скелетных мышц после кратковременного переедания у здоровых людей. Метаболизм 67, 26–30. DOI: 10.1016 / j.metabol.2016.10.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Там, К.С., Ковингтон, Дж. Д., Байпей, С., Чукалова, Ю., Бурк, Д., Йоханнсен, Д. Л. и др. (2014). Увеличение веса показывает резкое увеличение ремоделирования внеклеточного матрикса скелетных мышц. J. Clin. Эндокринол. Метаб. 99, 1749–1757. DOI: 10.1210 / jc.2013-4381
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Там, К. С., Пауэр, Дж. Э., Маркович, Т. П., Йи, К., Морш, М., МакЛеннан, С. В. и др. (2015). Влияние питания с высоким содержанием жиров на физическую функцию и внеклеточный матрикс скелетных мышц. Nutr. Диабет 5: e187. DOI: 10.1038 / nutd.2015.39
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Thorsteinsdóttir, S., Deries, M., Cachaço, A. S., and Bajanca, F. (2011). Размер внеклеточного матрикса развития скелетных мышц. Dev. Биол. 354, 191–207. DOI: 10.1016 / j.ydbio.2011.03.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тирни, М. Т., Громова, А., Сесилло, Ф. Б., Сала, Д., Спенле, К., Orend, G., et al. (2016). Автономное ремоделирование внеклеточного матрикса контролирует прогрессивную адаптацию регенеративной способности мышечных стволовых клеток во время развития. Cell Rep. 14, 1940–1952. DOI: 10.1016 / j.celrep.2016.01.072
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уэдзуми А., Фукада С., Ямамото Н., Такеда С. и Цучида К. (2010). Мезенхимальные предшественники, отличные от сателлитных клеток, способствуют образованию эктопических жировых клеток в скелетных мышцах. Нат. Cell Biol. 12, 143–152. DOI: 10.1038 / ncb2014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Улен М., Оксволд П., Фагерберг Л., Лундберг Э., Йонассон К., Форсберг М. и др. (2010). К основанному на знаниях Атласу белков человека. Нат. Biotechnol. 28, 1248–1250. DOI: 10.1038 / nbt1210-1248
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Урчиуоло, А., Куарта, М., Морбидони, В., Гаттаццо, Ф., Молон С., Грумати П. и др. (2013). Коллаген VI регулирует самообновление сателлитных клеток и регенерацию мышц. Нат. Commun. 4: 1964. DOI: 10.1038 / ncomms2964
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Vallecillo-García, P., Orgeur, M., Vom Hofe-Schneider, S., Stumm, J., Kappert, V., Ibrahim, D. M., et al. (2017). Odd skipped-related 1 идентифицирует популяцию эмбриональных фибро-адипогенных предшественников, регулирующих миогенез во время развития конечностей. Нат. Commun. 8: 1218.
Google Scholar
Вирджилио, К. М., Мартин, К. С., Пирс, С. М., и Блемкер, С. С. (2015). Многомасштабные модели скелетных мышц показывают комплексное влияние мышечной дистрофии на механику тканей и восприимчивость к повреждениям. Интерфейс Focus 5: 20140080. DOI: 10.1098 / rsfs.2014.0080
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Visse, R., and Nagase, H. (2003). Матричные металлопротеиназы и тканевые ингибиторы металлопротеиназ: структура, функция и биохимия. Circ. Res. 92, 827–839. DOI: 10.1161 / 01.RES.0000070112.80711.3D
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wang, D., Eraslan, B., Wieland, T., Hallström, B., Hopf, T., Zolg, D. P., et al. (2019). Атлас обилия протеома и транскриптомов 29 здоровых тканей человека. Мол. Syst. Биол. 15: e8503. DOI: 10.15252 / msb.20188503
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Весснер, Б., Либенштайнер, М., Nachbauer, W., and Csapo, R. (2019). Возрастной ответ внеклеточного матрикса скелетных мышц на упражнения с отягощениями: пилотное исследование. Eur. J. Sport Sci. 19, 354–364. DOI: 10.1080 / 17461391.2018.1526974
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уайт, Дж. П., Риси, Дж. М., Вашингтон, Т. А., Сато, С., Ле, М. Е., Дэвис, Дж. М. и др. (2009). Вызванное перегрузкой ремоделирование внеклеточного матрикса скелетных мышц и рост миофибрилл у мышей, лишенных IL-6. Acta Physiol. 197, 321–332. DOI: 10.1111 / j.1748-1716.2009.02029.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вильшут, К. Дж., Хаагсман, Х. П., и Ролен, Б. А. Дж. (2010). Компоненты внеклеточного матрикса определяют поведение стволовых клеток мышц свиньи. Exp. Cell Res. 316, 341–352. DOI: 10.1016 / j.yexcr.2009.10.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wood, L. K., Kayupov, E., Gumucio, J. P., Мендиас, К. Л., Клафлин, Д. Р., Брукс, С. В. (2014). Внутренняя жесткость внеклеточного матрикса в скелетных мышцах мышей увеличивается с возрастом. J. Appl. Physiol. 117, 363–369. DOI: 10.1152 / japplphysiol.00256.2014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Yucesoy, C.A., Maas, H., Koopman, B.H.F.J.M., Grootenboer, H.J., и Huijing, P.A. (2006). Механизмы, вызывающие влияние положения мышц на проксимально-дистальные различия мышечной силы при передаче внемышечной миофасциальной силы. Med. Англ. Phys. 28, 214–226. DOI: 10.1016 / j.medengphy.2005.06.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Заммит П.С. (2017). Функция миогенных регуляторных факторов Myf5, MyoD, Myogenin и MRF4 в скелетных мышцах, сателлитных клетках и регенеративном миогенезе. Семин. Cell Dev. Биол. 72, 19–32. DOI: 10.1016 / j.semcdb.2017.11.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжан, К., и Гао, Ю. (2012). Конечно-элементный анализ механики боковой передачи силы в одиночном мышечном волокне. J. Biomech. 45, 2001–2006. DOI: 10.1016 / j.jbiomech.2012.04.026
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Zhu, J., Li, Y., Shen, W., Qiao, C., Ambrosio, F., Lavasani, M., et al. (2007). Взаимосвязь между трансформирующим фактором роста-β1, миостатином и декорином: последствия для фиброза скелетных мышц. J. Biol. Chem. 282, 25852–25863.DOI: 10.1074 / jbc.M704146200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Zong, H., Bastie, C.C., Xu, J., Fassler, R., Campbell, K.P., Kurland, I.J., et al. (2009). Резистентность к инсулину у мышей с дефицитом бета1-рецептора интегрина, специфичного для поперечно-полосатых мышц. J. Biol. Chem. 284, 4679–4688. DOI: 10.1074 / jbc.M807408200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цюгель, М., Маганарис, К. Н., Вильке, Дж., Юркат-Ротт, К., Klingler, W., Wearing, S.C., et al. (2018). Исследование фасциальных тканей в спортивной медицине: от молекул до адаптации тканей, травм и диагностики: утверждение консенсуса. Br. J. Sports Med. 52: 1497. DOI: 10.1136 / bjsports-2018-099308
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Взаимосвязь между возрастом, ИМТ, положением головы и жесткостью и эластичностью поверхностных мышц шеи у взрослых женщин
Rantanen, T. et al. . Сила мышц и индекс массы тела как долгосрочные предикторы смертности у изначально здоровых мужчин. J Gerontol A Biol Sci Med Sci. 55 (3), M168–73 (2000).
CAS Статья Google Scholar
Уитто Дж. Биохимия соединительной ткани стареющей дермы. Возрастные изменения коллагена и эластина. Dermatol Clin. 4 , 433–46 (1986).
CAS Статья Google Scholar
Одетти, П.Р., Боргольо А. и Роланди Р. Возрастное увеличение флуоресценции коллагена в подкожной клетчатке человека. Метаболизм. 41 , 655–8 (1992).
CAS Статья Google Scholar
Родригес, А. Дж. Мл., Родригес, К. Дж., Кунья, А. К. и Джин, Ю. Количественный анализ коллагеновых и эластических волокон в поперечной фасции при прямой и прямой паховой грыже. Rev Hosp Clin Fac Med Sao Paulo. 57 , 265–70 (2002).
Артикул Google Scholar
Kragstrup, T. W., Kjaer, M. & Mackey, A. L. Структурные, биохимические, клеточные и функциональные изменения внеклеточного матрикса скелетных мышц при старении. Scand J Med Sci Sports. 21 , 749–57 (2011).
CAS Статья Google Scholar
Трапп, Т.Влияние старения и длительной разгрузки на структуру и функцию скелетных мышц человека. Appl Physiol Nutr Metab. 34 , 459–464 (2009).
CAS Статья Google Scholar
Хаус, Дж. М., Карритерс, Дж. А., Траппе, С. В. и Траппе, Т. А. Коллаген, сшивание и конечные продукты продвинутого гликирования в стареющих скелетных мышцах человека. J Appl Physiol (1985). 103 , 2068–76 (2007).
CAS Статья Google Scholar
Баррос, Э. М. и др. . Старение эластических и коллагеновых волокон шейных межостистых связок человека. Spine J. 2 , 57–62 (2002).
ADS MathSciNet Статья Google Scholar
Piasecki, M., Ireland, A., Jones, D. A. & McPhee, J. S. Зависимое от возраста ремоделирование двигательных единиц в мышцах конечностей человека. Биогеронтология. 17 , 485–96 (2016).
Артикул Google Scholar
Наричи, М. В. и Маффулли, Н. Саркопения: характеристики, механизмы и функциональное значение. Br Med Bull. 95 , 139–159 (2010).
CAS Статья Google Scholar
Мезнарич, М., Эржен, И., Карен, П., Цветко, Э.Влияние старения на состав тяжелой цепи миозина грудино-ключично-сосцевидной мышцы человека. Ann Anat. 28 (216), 95–32 (2017).
Google Scholar
Куа А. Б., Вильгельм К. П. и Майбах Х. И. Эластичные свойства кожи человека: зависимость от возраста, пола и анатомической области. Arch Dermatol Res. 282 , 283–8 (1990).
CAS Статья Google Scholar
Родригес, К. Дж. И Родригес, А. Дж. Мл. Сравнительное исследование старения системы эластичных волокон диафрагмы и прямых мышц живота у крыс. Braz J Med Biol Res. 33 , 1449–54 (2000).
CAS Статья Google Scholar
Alnaqeeb, M. A., Al Zaid, N. S. & Goldspink, G. Изменения соединительной ткани и физические свойства развивающихся и стареющих скелетных мышц. J Anat. 139 , 677–689 (1984).
CAS PubMed PubMed Central Google Scholar
Simons, D. G. & Mense, S. Понимание и измерение мышечного тонуса в связи с клинической болью в мышцах. Боль. 7 , 1–17 (1998).
Артикул Google Scholar
Баумгарт Э. Жесткость — неизвестный мир механической науки. Травма. 31 (Дополнение), S-B14–23 (2000).
Google Scholar
Гавронский Г. и др. . Оценка вязкоупругих параметров скелетных мышц юных триатлонистов. Physiol Measur. 28 , 625–637 (2007).
ADS Статья Google Scholar
Эби, С. Ф. и др. . Эластография пассивной жесткости скелетных мышц сдвиговой волной: влияние пола и возраста на протяжении всей взрослой жизни. Clin Biomech (Бристоль, Эйвон). 30 , 22–7 (2015).
Артикул Google Scholar
Домир, З. Дж., Маккалоу, М. Б., Чен, К. и Ан, К. Н. Возможность использования магнитно-резонансной эластографии для изучения влияния старения на модуль сдвига скелетных мышц. J Appl Biomech. 25 , 93–7 (2009).
Артикул Google Scholar
Kocur, P. et al. . Влияние старения на механические свойства грудино-ключично-сосцевидной и трапециевидной мышц при переходе из положения лежа в положение сидя — поперечное исследование. Arch Gerontol Geriatr. 70 , 14–18 (2017).
Артикул Google Scholar
Дитч А.М., Кларк Х.М., Штайнер Дж. Н. и Соломон Н. П. Влияние возраста, пола и положения тела на тонус орофациальных мышц у здоровых взрослых. J Speech Lang Hear Res. 58 , 1145–50 (2015).
Артикул Google Scholar
Наричи, М. В., Маффулли, Н. и Маганарис, К. Н. Старение мышц и сухожилий человека. Disabil Rehabil. 30 , 1548–54 (2008).
Артикул Google Scholar
Янссен, И., Хеймсфилд, С. Б., Ван, З. М., Росс, Р.Масса и распределение скелетных мышц у 468 мужчин и женщин в возрасте 18–88 лет. J Appl Physiol (1985). 89 , 81–8 (2000).
CAS Статья Google Scholar
Манини, Т. М. и Кларк, Б. С. Динапения и старение: обновление. J Gerontol A Biol Sci Med Sci. 67 , 28–40 (2012).
Артикул Google Scholar
Скелетно-мышечные изменения с возрастом: клиническое значение Кэрол Б. Льюис, PT, GCS, MSG, MPA, PhD Шари Келлемс, ATC inLewis Red. Старение: проблема здравоохранения, 4-е издание Philadeplhia Davies Company. 14–124 (2002).
Park, M. S. et al. . Возрастные изменения в сагиттальном диапазоне движений и выравнивании шейки матки. Глобальный. Spine J. 4 , 151–156 (2014).
Google Scholar
Юкава, Ю. и др. . Возрастные изменения костной анатомии, выравнивания и диапазона движений шейного отдела позвоночника. Часть I. Радиографические данные более 1200 бессимптомных субъектов. Eur Spine J. 21 (8), 1492–8 (2012).
Артикул Google Scholar
Рейн, С. и Туми, Л. Т. Вариации положения головы и плеч у 160 бессимптомных женщин и мужчин. Arch Phys Med Rehab. 78 , 1215–1223 (1997).
CAS Статья Google Scholar
Баумгартнер Р. Н. Состав тела в здоровом старении. Ann N Y Acad Sci. 904 , 437–448 (2000).
ADS CAS Статья Google Scholar
Джексон, А.С., Янссен, И., Суи, X., Черч, Т.С. и Блэр, С.Н. Продольные изменения в составе тела, связанные со здоровым старением: мужчины в возрасте 20–96 лет. Br J Nutr. 107 (7), 1085–91 (2012).
CAS Статья Google Scholar
Линдл Р. С. и др. . Возрастные и гендерные сравнения мышечной силы 654 женщин и мужчин в возрасте 20–93 лет. J Appl Physiol (1985). 83 , 1581–7 (1997).
CAS Статья Google Scholar
Галлахер Д. и др. . Насколько полезен индекс массы тела для сравнения ожирения по возрасту, полу и этническим группам? Am J Epidemiol. 143 , 228–239 (1996).
CAS Статья Google Scholar
Taylor-Piliae, R.E. et al. . Подтверждение результатов нового краткого обследования физической активности среди мужчин и женщин в возрасте 60–69 лет. Am J Epidemiol. 164 , 598–606 (2006).
Артикул Google Scholar
Мистерска, Э., Янковски, Р. и Гловацки, М. Межкультурная адаптация Индекса инвалидности шеи и Копенгагенской шкалы функциональной инвалидности шеи для пациентов с болью в шее из-за дегенеративных и дископатических расстройств. Психометрические свойства польских версий. BMC Musculoskelet Disord. 29 , 12–84 (2011).
Google Scholar
Шагаег, Ф., Ахмади, А., Маруфи, Н., Саррафзаде, Дж.Оценка положения головы вперед в положении сидя и стоя. Eur Spine J. 25 , 3577–3582 (2016).
Артикул Google Scholar
Миотон, П. Р. О. www.myoton.com/technology (2018).
Agyapong-Badua, S., Warnera, M., Dinesh, S. & Stokes, M. Измерение влияния старения на мышечный тонус и механические свойства прямых мышц бедра и двуглавой мышцы плеча у здоровых мужчин и женщин с использованием новое портативное миометрическое устройство. Arch Geront Geriatr. 62 , 59–67 (2016).
Артикул Google Scholar
Uthaikhup, S. & Jull, G. Показатели в тесте на черепно-шейное сгибание изменяются у пожилых людей. Man Ther. 14 , 475–9 (2009).
Артикул Google Scholar
Оатис, К. Кинезиология. Механика и патомеханика движения человека.Филадельфия: Липпинкотт Уильямс и Уилкинс. 480–492 (2004).
Валенсуэла, С. и др. . Влияет ли осанка головы на положение подъязычной кости и электромиографическую активность грудино-ключично-сосцевидной мышцы у молодых взрослых? Cranio. 23 , 204–11 (2005).
Артикул Google Scholar
Майерс, Т. Слово по фасции. В анатомии поезда.Миофасциальный для мануальной терапии и терапии движением . 2-е издание Курчил Ливингстон Эльзевьер. 9–51 (2009).
Пурслоу П. П. Структура и функциональное значение изменений соединительной ткани внутри мышцы. Comp Biochem Physiol A Mol Integr Physiol. 133 , 947–66 (2002).
Артикул Google Scholar
Госселин, Л. Э., Адамс, К., Коттер, Т. А., Маккормик, Р.J. & Thomas, Д. П. Влияние физической тренировки на пассивной скованности в опорно-двигательном аппарате скелетных мышц: роль внеклеточного матрикса. J Appl Physiol (1985). 85 , 1011–6 (1998).
CAS Статья Google Scholar
Rodriguez, H. et al. . Гистологическое описание взаимодействия мышечных волокон и соединительной ткани фасции трапециевидной мышцы человека. Внутр.J. Morphol. 29 , 299–303 (2011).
Артикул Google Scholar
Кади Ф. и Торнелл Л. Э. Тренировка влияет на фенотип тяжелой цепи миозина в трапециевидной мышце у женщин — гистохимия и клеточная биология. Гистохимия. 112 , 73 (1999).
CAS Статья Google Scholar
Цветко Э., Карен П.& Eržen, I. Состав тяжелой цепи миозина грудино-ключично-сосцевидной мышцы человека. Ann Anat. 194 , 467–72 (2012).
CAS Статья Google Scholar
Mutungi, G. & Ranatunga, K. W. Вязкие, вязкоупругие и эластичные характеристики покоящихся быстрых и медленных мышечных волокон млекопитающих (крыс). J. Physiol. 496 , 827–836 (1996).
CAS Статья Google Scholar
Ди Ангелантонио, Э. и др. . Индекс массы тела и смертность от всех причин: метаанализ данных отдельных участников 239 проспективных исследований на четырех континентах. Ланцет. 388 , 776–86 (2016).
Артикул Google Scholar
Jian, D. W., Wang, T. G. и Wang, Y. C. Жесткость мышц шеи, количественно определенная с помощью соноэластографии, коррелирует с индексом массы тела и симптомами хронической боли в шее. Ультразвук Med Biol. 39 , 1356–61 (2013).
Артикул Google Scholar
Кук, З., Кирк, С., Лоуренсон, С. и Сэндфорд, С. Использование ИМТ в оценке недостаточности питания у пожилых людей: размышления о практике. Proc Nutr Soc. 64 , 313–7 (2005).
CAS Статья Google Scholar
Moghadam, R.Э., Рахнама, Л., Карими, Н., Амири, М. и Рахнама, М. Ультрасонографическое исследование поперечного сечения глубоких мышц-сгибателей шеи при прямом и нормальном положении головы. J Bodyw Mov Ther. 22 , 643–647 (2017).
Артикул Google Scholar
Goodarzi, F., Rahnama, L., Karimi, N., Baghi, R. & Jaberzadeh, S. Влияние прямой позы головы на толщину мышц разгибателей шеи: ультразвуковое исследование. J Manipulative Physiol Ther. 41 , 34–41 (2018).
Артикул Google Scholar
Вагнер, К. Х., Камерон-Смит, Д., Весснер, Б. и Францке, Б. Биомаркеры старения: от функции к молекулярной биологии. Питательные вещества. 2 , 8 (2016).
Google Scholar
Francis, P. et al. . Измерение здоровья мышц при старении. Биогеронтология. 18 , 901–911 (2017).
Артикул Google Scholar
Фолкнер, Дж. А., Ларкин, Л. М., Клафлин, Д. Р. и Брукс, С. В. Возрастные изменения структуры и функции скелетных мышц. Clin Exp Pharmacol Physiol. 34 , 1091–6 (2007).
CAS Статья Google Scholar
Гайдосик Р.L. Пассивная растяжимость скелетных мышц: обзор литературы с клиническими последствиями. Clin Biomech (Бристоль, Эйвон). 16 , 87–101 (2001).
CAS Статья Google Scholar
Quek, J., Pua, Y.H., Clark, R.A. & Bryant, A.L. Влияние грудного кифоза и положения головы вперед на диапазон движений шейки матки у взрослых. Man Ther. 18 , 65–71 (2013).
Артикул Google Scholar
Канг, Дж. Х. и др. . Влияние переднего положения головы на постуральное равновесие у работника, долгое время работающего на компьютере. Ann Rehabil Med. 36 , 98–104 (2012).
Артикул Google Scholar
Хури В., Кардинал Э. и Брассард П. Атрофия и жировая инфильтрация надостной мышцы: сонография в сравнении с МРТ. AJR Am J Roentgenol. 19 , 1105–11 (2008).
Артикул Google Scholar
Паван П. Г., Стекко А., Стерн Р. и Стекко К. Болезненные соединения: уплотнение в сравнении с фиброзом фасции. Curr Pain Headache Rep. 18 (8), 441 (2014).
Артикул Google Scholar
Причинные факторы изменения состава газа внутри упаковок
Как хорошо известно, кислород, водяной пар и т. Д. Являются причинными факторами порчи продуктов питания и лекарств.Следовательно, устранение ресурсов кислорода и водяного пара и контроль состава газов внутри упаковки может эффективно продлить гарантийный срок продуктов и улучшить качество продуктов. Поэтому широкое распространение получили новые методы упаковки с использованием различных барьерных упаковочных материалов, такие как MAP, CAP и вакуумная упаковка. Однако даже при контроле состава газов и применении материалов с высокими барьерными свойствами для предотвращения проникновения кислорода и водяного пара фактический эффект упаковки все еще сильно отличается от предполагаемого результата.В этой статье мы подробно расскажем о причинных факторах изменения состава газов внутри упаковок и о методах их испытаний.
1. Анализ газа в упаковках
По завершении упаковки состав газов внутри упаковок будет временно стабильным. Но по мере того, как период хранения продолжается, объем кислорода или других газов (которые чувствительны к продуктам) может измениться, что в дальнейшем изменит состав газов внутри упаковки и повлияет на качество продукта.Анализ показывает, что увеличение содержания кислорода и водяного пара внутри упаковки в основном вызвано следующими факторами.
Прежде всего, это газы извне упаковки. Газы могут проходить через упаковку двумя способами: проникновением и утечкой. Самая большая разница между ними — это объем проникающих газов и место проникновения. Проникновение означает, что газы проходят через упаковку со стороны высокой концентрации в сторону низкой концентрации. Скорость проникновения в основном определяется барьерными свойствами упаковки.Эффективно решить эту проблему могут материалы с высокими показателями барьерных свойств. Но под утечкой понимается то, что газы проходят через упаковку через ее трещины, крошечные отверстия или неплотное соединение между двумя материалами. Такая утечка, скорее всего, обнаруживается при горячем запечатывании упаковок или вокруг крышек контейнеров. В пакете утечка и просачивание существуют одновременно. Таким образом, при проведении тестов следует обращать внимание на общий эффект от них обоих.
Второй фактор — это газы внутри упаковок, и такие газы включают оставшиеся газы и адсорбированные газы.Остающиеся газы — это газы (например, кислород), которые остаются внутри упаковки после завершения процесса упаковки. Хотя в MAP, CAP и вакуумной упаковке есть процесс замены газов и дегазации (особенно до кислорода), точность работы и время весьма ограничены, эффект дегазации неудовлетворителен. Даже так называемая «вакуумная упаковка» просто пытается уменьшить объем газов, но не полностью дегазировать. Адсорбированные газы также важны, потому что, кроме металлов и стекла, другие материалы обладают свойством адсорбировать газы.Объем адсорбированных газов часто зависит от структуры материала и периода контакта между материалом и газами. Однако адсорбция не равна абсорбции. Если происходит изменение среды, окружающей упаковку, и концентрация газов внутри упаковки снижается до определенного уровня, адсорбированные газы будут высвобождаться.
2. Методы испытаний
Хотя состав газов внутри упаковки меняется очень медленно, такое изменение все же повлияет на гарантийный срок продукта.Производители использовали множество средств для контроля газов, чтобы замедлить увеличение содержания кислорода и водяного пара внутри упаковки. Например, многослойные барьерные материалы для улучшения барьерных свойств упаковок; размещение осушителя, абсорбента или поглотителя кислорода для поглощения оставшихся газов и проникающих газов снаружи. Но каков эффект от этих мер? Требуется проанализировать пакет. Что касается каждого причинного фактора изменения состава газов, ниже приводится введение в соответствующие методы испытаний.
2.1 Тест на барьерные свойства
Тест на барьерные свойства очень важен для упаковочных материалов с высокими характеристиками барьерных свойств. Раньше можно было проверить только барьерные свойства пленки или хлопьев. Однако в реальной упаковке мы часто видим контейнеры и пакеты, поэтому данные испытаний материалов на барьерные свойства могут быть только справочными при выборе материалов. Эти данные не могут указать на окончательные барьерные свойства упаковок, поскольку толщина вокруг упаковок неодинакова, а некоторые свойства материалов могут измениться в процессе производства.Поэтому расчетный результат часто отличается от фактического результата теста. Исходя из такой ситуации, следует проверять не только барьерные свойства упаковочного материала, но также барьерные свойства конечной упаковки. Правильный выбор основан на данных обоих тестов. В настоящее время ведется разработка тестов на проникновение кислорода и водяного пара, и соответствующие испытательные приборы уже представлены на рынке. Эти инструменты вполне могут удовлетворить потребность в тестировании барьерных свойств целых упаковок.(Фото 1 — это прибор для измерения проницаемости кислорода в пленке / контейнере PERME серии TOY-C1 с использованием метода равного давления, который производится Labthink Languang; Фото 2 — это прибор для электролизного тестера проницаемости водяного пара TSY-W3 серии PERME) TOY-C1 Метод равного давления Пленка / контейнер для определения кислородопроницаемости
Фото 2. PERMETM TSY-W3 Электролизный тестер паропроницаемости для воды
2.2 Тест на герметичность
Тест на герметичность может указать на возможную утечку упаковка.Если воздух или жидкость попадут через упаковки через определенную точку утечки, барьерные свойства материалов упаковки полностью утратят свою функцию, и содержащиеся в ней продукты очень быстро ухудшатся. Соответствующий индекс герметичности отличается для каждой конкретной формы упаковки. Что касается пакетов с горячим запечатыванием, операция горячего запечатывания часто может снизить механическую прочность упаковок. Поэтому горячие запечатываемые детали обычно являются слабым местом герметичности мягкой упаковки. Более того, поскольку для герметизации мягких упаковок требуется давление, давление не распределяется равномерно на всю упаковку.Самая первая точка утечки — это часто та часть, которая испытывает наименьшее давление. В результате для объективной оценки необходимо проверить герметичность всей упаковки. Повышение давления внутри упаковок является основным методом проверки герметичности целых мягких упаковок. Когда мы проверяем герметичность мягких упаковок, нам необходимо смоделировать реальную ситуацию, когда они испытывают давление, создавая разницу давлений внутри и снаружи упаковок. Существует два способа выполнить такое моделирование: метод положительного давления (для надувания упаковки и создания большего внутреннего давления, например, продукт Labthink Languang, серия PARAM тестеров утечки и прочности уплотнения LSSD-01) и метод отрицательного давления (для создания вакуумная среда и снижение давления вне мешка, такие продукты, как Labthink Languang PARAM series MFY-01 Tightness Tester).Что касается контейнеров с винтом, то на герметичность, скорее всего, влияют стыки между крышками и бутылками. Такой тест может быть проведен на продуктах Labthink Languang серии PARAM LSSD -01 Leak and Seal Strength Tester.
2.3 Анализ газов в свободном пространстве
Процесс упаковки завершен, но это не означает, что мы не должны больше обращать на него внимание; внутри упаковки еще много газов. После завершения наполнения очень сложно контролировать и изменять состав газов до тех пор, пока упаковка не будет открыта.Упаковочные материалы с барьерными свойствами могут только замедлять проникновение газов, но не удалять газы внутри упаковки (за исключением тех случаев, когда применяется технология деоксигенации). Если объем оставшихся газов превышает предел максимальной концентрации, то ни один защитный материал и технология упаковки не могут удовлетворить требования гарантии на продукт. Поэтому нам необходимо проверить оставшиеся газы внутри упаковок и соответствующим образом скорректировать методы упаковки.
Как оставшиеся газы, так и адсорбированные газы могут быть проверены путем проверки газов в верхнем пространстве — проверки состава газов, собирающихся в верхней части упаковок.Единственная разница — время теста. Если нужно проверить оставшиеся газы, тесты следует проводить сразу после завершения процесса упаковки. Если мы хотим проверить адсорбированные газы, мы должны пройти тест в течение определенного периода времени после процесса упаковки и проводить тесты несколько раз с течением времени. Для всестороннего анализа необходимо построить кривую концентрации газов, поскольку выделение газов также является медленным процессом. Такой тест может быть проведен с помощью газоанализатора HGA-01 серии PARAM производства Labthink Languang.Конечно, мы также можем снизить концентрацию определенных газов в среде упаковки, чтобы ускорить выделение адсорбированных газов.
3. Резюме
В итоге, хотя изменения состава газов внутри упаковок избежать невозможно, мы можем всесторонне проанализировать продукт и спрогнозировать его гарантийный срок, проверив барьерные свойства всей упаковки, герметичность упаковок и состав. смена газов в наддувном пространстве. Эти тесты также могут помочь нам выявить слабые места в конструкции нашего пакета и предоставить нам точные данные тестирования для улучшения структуры наших пакетов.
Жесткость, прочность и твердость в производстве металлов
Кажется, существует некоторая путаница в отношении разницы между прочностью и жесткостью в металле. В конце концов, разве прочный металл не жесткий, а жесткий металл — крепкий?
Хотя часто существует корреляция между прочностью и жесткостью металла, для производственных целей эти два термина действительно следует четко разделять.
Почему?
Чтобы ответить на этот вопрос, вот несколько различий в значении жесткости и прочности металлических форм:
Определение жесткости
Жесткость — это жесткость любого объекта или материала.Предметы с высокой жесткостью будут сопротивляться изменению формы под действием физической силы.
Например, рыхлая влажная глина имеет низкую жесткость и меняет форму всего лишь под давлением в несколько фунтов. По жесткости алюминий значительно жестче влажной глины. Для сравнения, стальные сплавы обладают очень высокой жесткостью, что позволяет им выдерживать давление в тысячи фунтов на квадратный дюйм (PSI) без деформации поверхности металла.
Способность выдерживать давление, не сгибаясь, звучит так, будто ее следует использовать в качестве основы прочности металлического предмета, такого как проволочная корзина, но это не так.
На это тоже есть веская причина.
Жесткость зависит от модуля упругости, также известного как модуль Юнга. Хотя прочность может варьироваться от марки к марке, модуль Юнга постоянен для любого данного металла и не зависит от внешних факторов стресса, таких как термообработка, обработка или холодная обработка.
Проще говоря, когда дело доходит до прочности по сравнению с жесткостью, прочность металла может изменяться по всему объекту, но его жесткость остается постоянной.
Определение силы
Механическую прочность можно определить как меру напряжения, которое может быть приложено к материалу до того, как он деформируется или сломается.Это может сбивать с толку, поскольку сопротивление физической деформации является ключевой частью определения как прочности, так и жесткости.
Одно из ключевых отличий заключается в использовании слова «постоянный». Прочный материал может изгибаться под действием силы, которая ниже его максимального предела текучести, но он вернется к своей первоначальной форме, как только эта сила будет устранена. Это означает, что прочный материал может гнуться, не ломаясь, и вернуться к своей нормальной форме.
Еще одно ключевое отличие состоит в том, что прочность материала зависит от его химического состава и любых термических обработок, которым он подвергается при преобразовании в размер или форму.Проще говоря, прочность металла может варьироваться в зависимости от процессов, которым подвергался металл.
Определение твердости
В то время как первые два свойства определялись величиной напряжения или силы, которые металл может принять перед физическим воздействием, твердость измеряет сопротивление металла деформации поверхности. Обычно это относится к локализованному участку поверхности.
Например, если вы ударите молотком по поверхности куска дерева, вы заметите вмятину или отпечаток того места, где произошел удар.Это показатель твердости древесины. На противоположном конце спектра, если вы проделали то же самое с толстой стальной балкой и на поверхности не было визуальной деформации, вы могли бы сказать, что балка была намного тверже дерева.
Что означает разница для моих материалов?
Как указывалось ранее, часто существует сильная корреляция между прочностью, твердостью и жесткостью. Но разница может быть большой, особенно при производстве металлических материалов, которые будут подвергаться воздействию множества внешних сил.
Например, чрезвычайно жесткий материал также может быть очень хрупким, склонным к разрушению при превышении порогового значения давления. С другой стороны, прочные материалы имеют тенденцию изгибаться до того, как они сломаются, и отскакивают после снятия нагрузки, если только указанная нагрузка не превышает предел текучести материала. И, в зависимости от твердости материала, независимо от прочности или жесткости, он может быть поврежден или не поврежден ударом другого объекта.
Итак, когда вы спрашиваете, насколько прочны изделия изготовителя металлических форм на заказ, убедитесь, что они знают разницу между прочностью, жесткостью и твердостью.
Позвольте Marlin Steel направлять ваше производство металла
Вам нужны металлические изделия, отвечающие вашим требованиям по прочности, твердости или жесткости? Команда экспертов Marlin Steel имеет обширный опыт работы с широким спектром материалов, таких как обычная, нержавеющая, оцинкованная сталь, алюминий, титан и множество других сплавов, которые различаются по составу.