ПЦР. Флороценоз бактериальный вагиноз (количественно) (4 показателя: ДНК Bacteria, ДНК Lactobacillus spp, ДНК Gardnerella vaginalis, ДНК Atopobium vaginae)
Срок выполнения, дней: 1-2
Код исследования: Р22
Бактериальный вагиноз состояние, при котором лактофлора (нормальные бактерии, образующие защитную биопленку и кислую pH< 4,5) полностью или частично замещается факультативно — анаэробными бактериями.Gardnerella vag.и Atopobium vag. – ключевые микроорганизмы, ответственные за развитие бактериального вагиноза.
Бактериальный вагиноз с современных позиций рассматривается как не воспалительный инфекционный синдром, характеризующийся комплексом качественных и количественных изменений в микрофлоре влагалища. Основное звено патогенеза БВ -образование биопленки, при БВ она состоит: 60-90 % G.vaginalis, 1-40 % A.
 vaginae, 1-15 % другие бактерии. При этом содержание лактобактерий снижается до 0.1-0.001 %
vaginae, 1-15 % другие бактерии. При этом содержание лактобактерий снижается до 0.1-0.001 %Флороценоз Бактериальный вагиноз позволяет с высокой аналитической чувствительностью и специфичностью поставить диагноз бактериальный вагиноз, с количественным подсчетом основных маркеров, благодаря специальному программному обеспечению. A.vaginaeне чувствителен к метронидозолу и чувствителен к клиндамицину.
Подготовка пациента:
Перед исследованием в течение 7 дней не проводить вагинальных процедур (не ставить свечи, не использовать антисептики и т. д.).
Материал: Отделяемое заднего свода влагалища.
Стабильность пробы: При t 6-8° C в течение 1 недели, и длительно при замораживании.
Метод: ПЦР количественный анализ.
Анализатор: RotorGene.
Тест – система: АмплиСенс.
Референсные значения (норма):
ДНК Lactobacillus spp 105– 107 ГЭ/мл;
ДНК Gardnerella vaginalis < 5 ·103
ДНК Atopobium vaginae < 5 ·103 ГЭ/мл;
Соответствут нормофлоре – «нормоценоз».

Основные показания к назначению анализа:
1. Наличие специфических выделений, увеличение ph >4.5, наличие «ключевых клеток» при микроскопии.
Интерпретация результатов: Расчет ведется с помощью программного обеспечения и выдается в виде заключения:
lgLacto-lg(Gv+Av)
| <0.5 | Полученные параметры соответствуют Бактериальному вагинозу |
| >1 | На основании полученных параметров Бактериальный вагиноз не выявлен |
| 0.5-1 | Полученные параметры соответствуют Промежуточному типу микрофлоры |
Положительный результат:
ДНК Lactobacillus spp <105 ГЭ/мл;
ДНК Gardnerella vaginalis > 5 ·105 ГЭ/мл;
ДНКAtopobium vaginae > 5 ·105 ГЭ/мл.
Отрицательный результат:
ДНК Lactobacillus spp 105– 107 ГЭ/мл;
ДНК Gardnerella vaginalis < 5 ·103 ГЭ/мл;
ДНК Atopobium vaginae < 5 ·103 ГЭ/мл. <0.5 Полученные параметры соответствуют
Бактериальному вагинозу >1 На основании полученных параметров
Бактериальный вагиноз не выявлен 0.5-1 Полученные параметры соответствуют
Промежуточному типу микрофлоры
Что нужно знать о бактериальном вагинозе
Многие женщины сталкиваются с симптомами влагалищного дисбиоза. Обильные выделения, молочно-серого цвета и с неприятным запахом часто путают с «молочницей» и проводят «соответствующее» самолечение.
В большинстве случаев, такая стратегия быстрее навредит, чем поможет, поскольку причиной может быть бактериальный вагиноз (БВ).
Что такое баквагиноз
Согласно официальному определению, это невоспалительное заболевание влагалища, характеризующееся резким снижением или полным отсутствием лактофлоры и её заменой на анаэробные бактерии, среди которых наиболее распространены Gardnerella vaginalis (гарднерелла) и Atopobium vaginae (атопобиум).
У здоровых женщин репродуктивного возраста влагалищный микробиом на 95% представлен лактобактериями. Их способность к кислотообразованию (молочная кислота) и продукции перекиси водорода способствует поддержанию кислого pH и подавлению роста остальных микроорганизмов.
Оставшиеся 5% микрофлоры занимают более 300 видов аэробных и анаэробных микроорганизмов, среди которых:
- грамположительные палочки — Corynebacterium spp., Eubacterium spp., Bifidobacterium spp., Propionibacterium spp., Clostridium spp.;
- грамотрицательные палочки (14-55%) — Bacteroides spp., Fusobacterium spp., Prevotella spp., Porphyromonas spp., Leptotrichia spp.;
- грамположительные кокки (30-80%) — Peptostreptococcus spp., Peptococcus spp., Staphylococcus spp., Micrococcus varians, Enterococcus spp., Streptococcus viridans и др.
- грамотрицательные кокки — Veilonella spp., Acidominococcus fermentans, непатогенные Neisseria spp.,
- энтеробактерии — Echerihia coli, Klebsiella spp.
 , Enterobacter spp., Citrobacter spp. и др.
, Enterobacter spp., Citrobacter spp. и др.
 , Enterobacter spp., Citrobacter spp. и др.
, Enterobacter spp., Citrobacter spp. и др.Все это многообразие бактерий является частью нормального «женского» биоценоза, не вызывая симптомов и не требуя лечения. Однако это правомерно до тех пор, пока их численность лактобацилл не начинает снижаться, а остальных — расти.
Факторы риска и симптомы
Почему происходит такой «сдвиг» — достоверно пока не известно. Однако к факторам риска относят:- ранее перенесенные воспалительные заболевания половых путей,
- длительный и бесконтрольный прием антибиотиков,
- гормональные нарушения (расстройства менструального цикла),
- длительное применение ВМС,
- прием оральных контрацептивов.
Колонизация влагалища микрофлорой, связанной с баквагинозом, особенно Gardnerella vaginalis и Atopobium vaginae, сопровождается:
- сдвигом pH в щелочную сторону (>4,5),
- появлением обильных (иногда пенистых) выделений молочно-серого цвета
- появлением характерного «рыбного» запаха, усиливающегося при защелачивании среды (половой контакт, менструация).

Отека и покраснения обычно не наблюдается, что является отличительным признаком от банального вагинита и кандидоза влагалища («молочницы»).
Диагностика
Самым простым, но косвенным, признаком нарушения влагалищной микрофлоры служит повышение pH >4,5. Для проведения такого «анализа» требуются всего лишь тест-полоски для измерения рН, доступные к покупке в любой аптеке. Однако, чтобы выявить возбудителя и начать адекватную терапию такой диагностики, конечно, недостаточно.
Современным и высокоточным методом диагностики влагалищных дисбиозов служит комплексное исследование «Флороценоз».
Этот тест разработан научными специалистами совместно с практикующими клиницистами. И предназначен для определения состава и соотношения нормальной и условно-патогенной флоры влагалища методом ПЦР.
Исследование представлено в 3-х вариантах для различных клинических ситуаций:
1. Флороценоз – бактериальный вагиноз
Название теста «говорит само за себя», а в результате будет отражено количество:
- всех обнаруженных бактерий,
- ДНК Lactobacillus spp. ,
- ДНК Atopobium vaginae,
- ДНК Gardnerella vaginalis.
 ,
,2. В случае, когда есть подозрения на нарушения более «широкого спектра», к применению рекомендован «Флороценоз», включающий подсчет:
- ДНК всех бактерий,
- ДНК Lactobacillus spp.,
- ДНК возбудителей бактериального вагиноза
- Gardnerella vaginalis
- и Atopobium vaginae,
- ДНКвозбудителейаэробноговагинита
- Enterobacteriaceae,
- Staphylococcus spp.,
- Streptococcus spp.,
- ДНК условно-патогенных микоплазм
- Ureaplasma urealyticum,
- Ureaplasma parvum,
- Mycoplasma hominis,
- ДНК возбудителей кандидоза («молочницы»)
- Candida albicans,
- Candida glabrata,
- Candida krusei
- Candida paraps./ tropicalis.
3. При необходимости в дополнительной диагностике ИППП – подходит «Флороценоз – комплексное исследование», включающий в себя подсчет «классического состава» бактерий + выявление ДНК «виновников» ИППП:
При необходимости в дополнительной диагностике ИППП – подходит «Флороценоз – комплексное исследование», включающий в себя подсчет «классического состава» бактерий + выявление ДНК «виновников» ИППП:
- Neisseria gonorrhoeae (гонокок)
- Chlamydia trachomatis (хламидия)
- Mycoplasma genitalium (микоплазма гениталиум)
- Trichomonas vaginalis (трихомонада)
Готовность анализов составляет 4-5 дней, а результат можно получить по электронной почте, в личном кабинете на сайте или в любом удобном отделении KDL.
Поделиться статьей:
Остались вопросы?Флороценоз Бактериальный вагиноз (количественное определение ДНК Gardnerella vaginalis, Atopobium vaginae, Lactobacillus spp.
 , Bacteria)
, Bacteria)Флороценоз Бактериальный вагиноз – это комплексное исследование состава микрофлоры влагалища у женщин репродуктивного возраста (18 — 45 лет) для диагностики бактериального вагиноза. Исследование характеризует общую бактериальную массу, выявляет и дает количественную оценку нормальной микрофлоры и условно-патогенных микроорганизмов, маркеров бактериального вагиноза.
Условно-патогенные микроорганизмы могут в небольших количествах определяться в норме в пробах из влагалища у здоровых женщин. При неблагоприятных условиях и при увеличении концентраций условно-патогенные микроорганизмы могут вызывать дисбиозы и воспалительные заболевания мочеполовой системы, приводить к развитию осложнений.
Бактериальный вагиноз — это состояние, при котором нарушен баланс микрофлоры влагалища. При бактериальном вагинозе снижается доля нормальной микрофлоры (лактобактерий), а количество условно-патогенных бактерий значительно возрастает.
Исследование Флороценоз Бактериальный вагиноз проводится методом количественного ПЦР в режиме реального времени. ПЦР является прямым методом диагностики инфекций, он выявляет участки ДНК исследуемых микроорганизмов. Анализ является высокочувствительным, специфичным, автоматизированным и быстрым в исполнении.
В состав исследования Флороценоз Бактериальный вагиноз входят
- Bacteria – показывает общее количество бактерий.
- Gardnerella vaginalis – Гарднерелла вагиналис, условно-патогенный микроорганизм, маркер бактериального вагиноза.
- Lactobacillus spp. — Лактобактерии, нормальная микрофлора влагалища для женщин репродуктивного возраста.
- Atopobium vaginae – Атопобиум вагине, условно-патогенный микроорганизм, маркер бактериального вагиноза.
Исследование показано
- При признаках воспалительных заболеваний половых органов у женщин: если имеются выделения из влагалища, зуд, жжение половых органов, неприятный запах, дискомфорт, болезненность при половых контактах, нарушено мочеиспускание.
- Для исключения инфекций мочеполового тракта, которые протекают «скрыто», без выраженных признаков заболеваний.
- При планировании беременности.
- Для обследования перед оперативным лечением на половых органах и органах малого таза.
- Для контроля после проведенного лечения.

Результат исследования
Учёт и трактовка результатов реакции проводится автоматически с помощью программного обеспечения.
Проводится определение общего количества бактерий, количества нормальной микрофлоры и условно-патогенных микроорганизмов.
Абсолютная концентрация участков ДНК микроорганизмов рассчитывается в геномных эквивалентах в 1 мл биологического образца (ГЭ/мл).
На основании соотношений концентраций исследуемых микроорганизмов формируется заключение по проведенному исследованию. В заключении дается характеристика состоянию микрофлоры влагалища, сообщается о наличии или отсутствии бактериального вагиноза.
Материал для исследования
В качестве биологического материала для анализа используется соскоб из влагалища.
Так как ПЦР анализ является прямым методом определения возбудителей инфекций, то он обнаружит причину заболевания, только если микроорганизм будет содержаться в исследуемом материале.
Трактовку результата исследования проводит врач акушер-гинеколог, принимая во внимание данные осмотра, проявления заболевания, признаки наличия воспаления (увеличение числа лейкоцитов) по анализу урогенитального мазка на флору, результаты всех дополнительных обследований. Врач определяет необходимость и вид лечения.
| Код | Название | Материал | Срок исполнения | Цена | ||
Комплексное исследование на выявление возбудителей инфекций мочеполовой системы |
||||||
| 170001 | Комплексное исследование ИППП (Chlamydia trachomatis, Mycoplasma genitalium, Neisseria gonorrhoeae, Trichomonas vaginalis), количественное определение ДНК | соскоб из урогенитального тракта, моча |
2 р. д. д.
|
770,00р. | ||
| 170004 |
Урогенитальные инфекции у мужчин (N. gonorrhoeae/C. trachomatis/ M. genitalium/ T. vaginalis// U. parvum/urealyticum/M. hominis//C.albicans/glabrata/crusei), количественное определение ДНК |
соскоб из урогенитального тракта, моча | 2 р.д. | 1910,00р. | ||
| 170005 |
Урогенитальные инфекции у женщин (N. gonorrhoeae/ C. trachomatis/M. genitalium/ T. vaginalis//U. parvum/ urealyticum/M. hominis// C.albicans/glabrata/crusei// Бактериальный вагиноз), количественное определение ДНК |
Соскоб из урогенитального тракта | 3 р.д |
2310,00р.
|
||
| 000005 | Комплексное исследование ИППП (Chlamydia trachomatis, Mycoplasma genitalium, Neisseria gonorrhoeae,Trichomonas vaginalis), качественное определение рибосомальной РНК методом NASBA |
У мужчин: отделяемое уретры, моча. У женщин: отделяемое цервикального канала, влагалища |
2 р.д. | 5000,00р. | ||
| 140021 | Комплексное исследование на возбудителей кандидоза, гонореи, трихомоноза (Candida, Neisseria gonorrhoeae,Trichomonas vaginalis) | мазок из урогенитального тракта | 5 р.д. | 1620,00р. | ||
| 310001 |
Флороценоз (ДНК Candida albicans, ДНК Candida glabrata, ДНК Candida krusei, ДНК Candida parapsilosis, ДНК Candida tropicalis, ДНК Ureaplasma parvum, ДНК Ureaplasma urealyticum, ДНК Mycoplasma hominis, ДНК Cardnerella vaginalis, ДНК Atopobium vaginae, ДНК Enterobacteriaceae, ДНК Staphylococcus spp. , ДНК Streptococcus spp., ДНК Lactobacillus spp., ДНК Bacteria spp.) , ДНК Streptococcus spp., ДНК Lactobacillus spp., ДНК Bacteria spp.)
|
мазок из влагалища | 3 р.д. | 1555,00р. | ||
| 310002 | Флороценоз и Микроскопия (ДНК Candida albicans, ДНК Candida glabrata, ДНК Candida krusei, ДНК Candida parapsilosis, ДНК Candida tropicalis, ДНК Ureaplasma parvum, ДНК Ureaplasma urealyticum, ДНК Mycoplasma hominis, ДНК Cardnerella vaginalis, ДНК Atopobium vaginae, ДНК Enterobacteriaceae, ДНК Staphylococcus spp., ДНК Streptococcus spp., ДНК Lactobacillus spp., ДНК Bacteria spp.), Микроскопическое исследование мазка из 2 точек | Мазок из влагалища, мазок из влагалища и цервикального канала | 3 р.д. | 1840,00р. | ||
| 310003 |
Флороценоз и NCMT (ДНК Candida albicans, ДНК Candida glabrata, ДНК Candida krusei, ДНК Candida parapsilosis, ДНК Candida tropicalis, ДНК Ureaplasma parvum, ДНК Ureaplasma urealyticum, ДНК Mycoplasma hominis, ДНК Cardnerella vaginalis, ДНК Atopobium vaginae, ДНК Enterobacteriaceae, ДНК Staphylococcus spp. , ДНК Streptococcus spp., ДНК Lactobacillus spp., ДНК Bacteria spp., ДНК Neisseria gonorrhoeae, ДНК Chlamydia trachomatis, ДНК Mycoplasma genitalium, ДНК Trichomonas vaginalis) , ДНК Streptococcus spp., ДНК Lactobacillus spp., ДНК Bacteria spp., ДНК Neisseria gonorrhoeae, ДНК Chlamydia trachomatis, ДНК Mycoplasma genitalium, ДНК Trichomonas vaginalis)
|
Мазок из влагалища | 3 р.д. | 2150,00р. | ||
| 310004 | Флороценоз и NCMT и Микроскопия (ДНК Candida albicans, ДНК Candida glabrata, ДНК Candida krusei, ДНК Candida parapsilosis, ДНК Candida tropicalis, ДНК Ureaplasma parvum, ДНК Ureaplasma urealyticum, ДНК Mycoplasma hominis, ДНК Cardnerella vaginalis, ДНК Atopobium vaginae, ДНК Enterobacteriaceae, ДНК Staphylococcus spp., ДНК Streptococcus spp., ДНК Lactobacillus spp., ДНК Bacteria spp., ДНК Neisseria gonorrhoeae, ДНК Chlamydia trachomatis, ДНК Mycoplasma genitalium, ДНК Trichomonas vaginalis), Микроскопическое исследование мазка из 2 точек | Мазок из влагалища, мазок из влагалища и цервикального канала |
3 р. д. д.
|
2530,00р. | ||
Хламидии |
||||||
| 010001 | Хламидия (Chlamydia trachomatis), качественное определение ДНК | соскоб из урогенит. тракта, моча, синов. ж-ть, сперма, секрет простаты, мазок с конъюнктивы, мазок из ротоглотки | 3 р.д | 255,00р. | ||
| 010003 | Хламидия (Chlamydia trachomatis), количественное определение ДНК | соскоб из урогенит. тракта, моча | 3 р.д | 895,00р. | ||
| 041401 | Хламидия (Chlamydia trachomatis), полуколичественное определение антител класса IgG | кровь (сыворотка) |
2 р. |
595,00р. | ||
| 041402 | Хламидия (Chlamydia trachomatis), полуколичественное определение антител класса IgА | кровь (сыворотка) | 2 р.д. | 600,00р. | ||
| 041407 | Хламидия (Chlamydia trachomatis), качественное определение антител класса IgМ | кровь (сыворотка) | 2 р.д. | 580,00р. | ||
| 000001 | Хламидия (Chlamydia trachomatis), качественное определение рибосомальной РНК методом NASBA |
У мужчин: отделяемое уретры, моча. У женщин: отделяемое цервикального канала, влагалища |
2 р.д. | 1560,00р. | ||
Микоплазмы |
||||||
| 042001 | Уреаплазма (Ureaplasma urealyticum), качественное определение антител класса IgG | кровь (сыворотка) |
2 р. д. д.
|
595,00р. | ||
| 042002 | Уреаплазма (Ureaplasma urealyticum), качественное определение антител класса IgА | кровь (сыворотка) | 2 р.д. | 575,00р. | ||
| 042003 | Уреаплазма (Ureaplasma urealyticum), качественное определение антител класса IgМ | кровь (сыворотка) | 2 р.д. | 575,00р. | ||
| 140001 | Уреаплазма (U.urealyticum / U. parvum), посев с определением чувствительности к антибиотикам | соскоб, сперма, секрет простаты, моча |
7 р.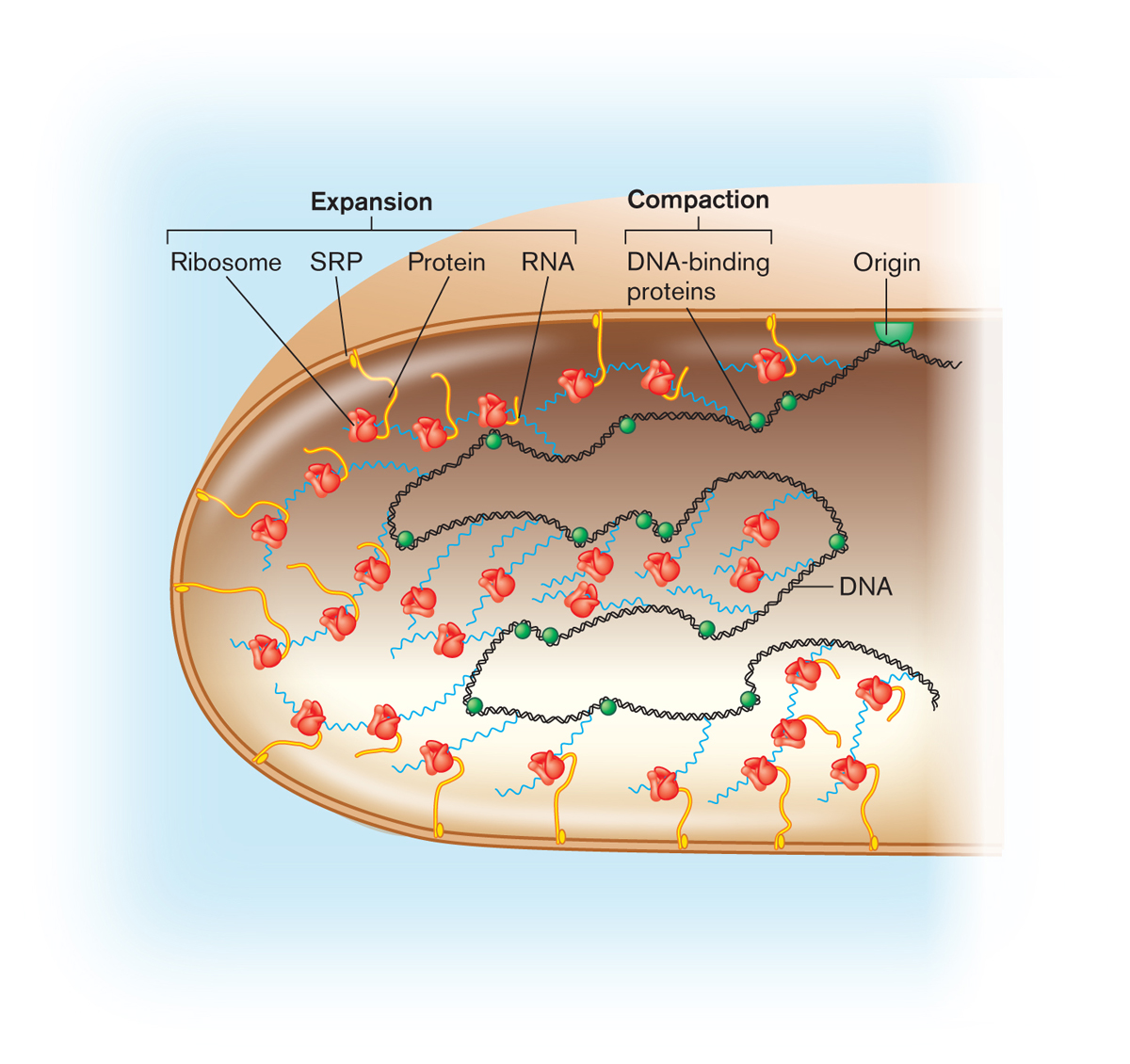 д д
|
860,00р. | ||
| 010107 | Типирование уреаплазмы (U.urealyticum / U. parvum), качественное определение ДНК | соскоб из урогенит. тракта, моча, секр. простаты, сперма | 3 р.д | 255,00р. | ||
| 010109 | Типирование уреаплазмы (U.urealyticum / U. parvum), количественное определение ДНК | соскоб из урогенит. тракта, моча | 3 р.д. | 565,00р. | ||
| 041503 | Микоплазма (Mycoplasma pneumoniae), количественное определение антител класса IgG | кровь (сыворотка) |
2 р. д. д.
|
650,00р. | ||
| 041504 | Микоплазма (Mycoplasma pneumoniae), количественное определение антител класса IgА | кровь (сыворотка) | 2 р.д. | 640,00р. | ||
| 041505 | Микоплазма (Mycoplasma pneumoniae), качественное определение антител класса IgМ | кровь (сыворотка) | 2 р.д. | 620,00р. | ||
| 010101 | Микоплазма (Mycoplasma hominis), качественное определение ДНК | соскоб из урогенит. тракта, моча, сперма, секрет простаты |
3 р. д д
|
255,00р. | ||
| 010104 | Микоплазма (Mycoplasma hominis), количественное определение ДНК | соскоб из урогенит. тракта, моча | 3 р.д. | 615,00р. | ||
| 010102 | Микоплазма (Mycoplasma genitalium), качественное определение ДНК | соскоб из урогенит. тракта, моча, сперма, секрет простаты | 3 р.д | 255,00р. | ||
| 010110 | Микоплазма (Mycoplasma genitalium), количественное определение ДНК | соскоб из урогенит. тракта, моча | 3 р.д | 895,00р. | ||
| 170002 | Микоплазмы, комплексное исследование (Ureaplasma urealyticum, Ureaplasma Parvum, Mycoplasma hominis), количественное определение ДНК |
соскоб из урогенит. тракта тракта
|
3 р.д. | 920,00р. | ||
| 140002 | Микоплазма (Mycoplasma hominis), посев с определением чувствительности к антибиотикам | соскоб, сперма, секрет простаты, моча | 7 р.д. | 860,00р. | ||
| 041501 | Микоплазма (Mycoplasma hominis), качественное определение антител класса IgG | кровь (сыворотка) | 2 р.д. | 540,00р. | ||
| 041502 | Микоплазма (Mycoplasma hominis), качественное определение антител класса IgМ | кровь (сыворотка) | 2 р.д. |
565,00р.
|
||
| 041506 | Микоплазма (Mycoplasma hominis), качественное определение антител класса IgА | кровь (сыворотка) | 2 р.д. | 645,00р. | ||
| 140036 | Диагностика микоплазменной инфекции, посев (Ureaplasma spp./Mycoplasma hominis), определение количества и чувствительности к антибиотикам | соскоб |
3 р.д. |
2900,00р. | ||
| 000003 | Микоплазма (Mycoplasma genitalium), качественное определение рибосомальной РНК методом NASBA |
У мужчин: отделяемое уретры, моча. У женщин: отделяемое цервикального канала, влагалища |
2 р. д. д.
|
1560,00р. | ||
|
Гарднерелла |
||||||
| 010201 | Гарднерелла (Gardnerella vaginalis), качественное определение ДНК | соскоб из урогенит. тракта | 3 р.д | 255,00р. | ||
| 170301 | Бактериальный вагиноз(Gardnerella vaginalis/ Lactobacillus sp/ Atopobium vaginae/ количество клеток), количественное определение ДНК | соскоб из жен. урогенитального тракта | 3 р.д. | 955,00р. | ||
Гонорея |
||||||
| 010401 | Возбудитель гонореи (Neisseria gonorrhoeae), качественное определение ДНК |
соскоб из урогенит. тракта, моча, секрет простаты, сперма тракта, моча, секрет простаты, сперма
|
3 р.д | 255,00р. | ||
| 010404 | Возбудитель гонореи (Neisseria gonorrhoeae), количественное определение ДНК | соскоб из урогенит. тракта, моча | 3 р.д | 895,00р. | ||
| 140014 | Возбудитель гонореи (Neisseria gonorrhoeae), посев с определением чувствительности к антибиотикам | мазок из урогенитального тракта | 5 р.д. | 960,00р. | ||
| 000002 | Возбудитель гонореи (Neisseria gonorrhoeae), качественное определение рибосомальной РНК методом NASBA |
У мужчин: отделяемое уретры, моча. У женщин: отделяемое цервикального канала, влагалища |
2 р. д. д.
|
1560,00р. | ||
Трихомонады |
||||||
| 020201 | Возбудитель трихомоноза (Trichomonas vaginalis), качественное определение ДНК | соскоб из урогенит. тракта, моча, секрет простаты, сперма | 3 р.д | 255,00р. | ||
| 020202 | Возбудитель трихомоноза (Trichomonas vaginalis), количественное определение ДНК | соскоб из урогенит. тракта, моча | 3 р.д | 955,00р. | ||
| 140020 | Возбудитель трихомоноза (Trichomonas vaginalis), посев | мазок из урогенитального тракта | 5 р.д. |
940,00р.
|
||
| 000004 | Возбудитель трихомоноза (Trichomonas vaginalis), качественное определение рибосомальной РНК методом NASBA |
У мужчин: отделяемое уретры, моча. У женщин: отделяемое цервикального канала, влагалища |
2 р.д. | 1560,00р. | ||
Вирус папилломы человека |
||||||
| 031203 | Вирус папилломы человека (Human papillomavirus, HPV) 6/11, качественное определение ДНК | соскоб из урогенитального тракта (цервик. rанал, шейка матки — у женщин; уретра, крайняя плоть — у мужчин) | 3 р.д | 255,00р. | ||
| 031201 | Вирус папилломы человека (Human papillomavirus, HPV) 16/18, качественное определение ДНК |
соскоб из урогенитального тракта (цервик. канал — у женщин; уретра, крайняя плоть — у мужчин) канал — у женщин; уретра, крайняя плоть — у мужчин)
|
3 р.д | 255,00р. | ||
| 031207 | Вирус папилломы человека (Human papillomavirus, HPV) 16/18, количественное определение ДНК | соскоб из урогенитального тракта (цервик. канал — у женщин; уретра, крайняя плоть — у мужчин) | 5 р.д | 640,00р. | ||
| 031208 | Вирус папилломы человека (Human papillomavirus, HPV) высокого канцерогенного риска (16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68),суммарное качественное определение ДНК | соскоб из урогенитального тракта (цервик. канал — у женщин; уретра, крайняя плоть — у мужчин) | 3 р.д | 530,00р. | ||
| 031209 | Вирус папилломы человека (Human papillomavirus, HPV) высокого канцерогенного риска (16, 18, 31, 33, 35, 39, 45, 51, 52,56, 58, 59), качественное определение ДНК с указанием типа вируса |
соскоб из урогенитального тракта (цервик. канал — у женщин; уретра, крайняя плоть — у мужчин) канал — у женщин; уретра, крайняя плоть — у мужчин)
|
7 р.д. | 860,00р. | ||
| 031206 | Вирус папилломы человека (Human papillomavirus, HPV) высокого канцерогенного риска (16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68), суммарное количественное определение ДНК | соскоб из урогенитального тракта (цервик. канал — у женщин; уретра, крайняя плоть — у мужчин) | 7 р.д. | 1025,00р. | ||
| 031210 | ВПЧ-тест расширенный (с определением количества и типа вируса) | цервикальный соскоб | 3-7 р.д. | 1540,00р. | ||
| 031211 | ВПЧ-ПАП-тест (комплекс тестов: ВПЧ расширенный с определением количества и типа вируса + ПАП-тест) | цервикальный соскоб (стекло+пробирка) |
3-7 р. д. д.
|
2500,00р. | ||
| 031213 | ВПЧ-тест расширенный жидкостный (с определением количества и типа вируса) | цервикальный соскоб жидкостный | 3-7 р.д. | 1610,00р. | ||
| 031212 | ВПЧ-ПАП-тест жидкостный (комплекс тестов:ВПЧ расширенный с опрделением количества и типа вируса +ПАП-тест) | цервикальный соскоб жидкостный | 3-7 р.д. | 3220,00р. | ||
| 031214 | ПАП-тест жидкостный | цервикальный соскоб жидкостный | 7 р.д. | 2340,00р. | ||
| 031218 | ВПЧ-тест (с определением количества и отдельным выявлением 16 и 18 типов вируса) | соскоб из цервикального канала, мазок из влагалища |
до 5 р. д. д. |
1090,00р. | ||
ПЦР Флороценоз-13 (Бак.вагиноз+Микоплазмы+Аэробы+Кандиды) — Клиника ЕВРОМЕД
ПЦР Флороценоз-13 (Бак.вагиноз+Микоплазмы+Аэробы+Кандиды)
1250 ₽
Комплексное исследование «Флороценоз» с включением NCMT — это универсальный тест для исследования бактериальной микрофлоры влагалища у женщин детородного возраста.
Дисбаланс вагинальной микрофлоры приводит к развитию целого комплекса осложнений, влияющих на репродуктивную функцию женщины (воспалительные заболевания органов малого таза, повышение риска развития поздних самопроизвольных выкидышей, преждевременных родов, постабортных эндометритов и др.).
Выявляются следующие микроорганизмы:
Нормальная и условно- патогенная флора:
- ДНК бактерий,
- Lactobacillus spp. ,
- Gardnerella vaginalis,
- Atopobium vaginae,
- Enterobacteriaceae,
- Staphylococcus spp.,
- Streptococcus spp.,
- Ureaplasma parvum,
- Ureaplasma urealyticum,
- Mycoplasma hominis,
- Candida albicans,
- ДНК Candida glabrata,
- Candida krusei,
- Candida paraps./ tropicalis.
 ,
,В норме для женщин репродуктивного возраста общая бактериальная масса (ДНК Bacteria) представлена, в основном, лактобактериями (концентрация ДНК Lactobacillus spp. не менее концентрации ДНК Bacteria). Концентрация облигатно-анаэробных бактерий (ДНК Gardnerella vaginalis и ДНК Atopobium vaginae) не должны превышать концентрацию лактобактерий (ДНК Lactobacillus spp.)
Показания к назначению анализа:
- Диагностика абсолютных патогенов: качественное определение хламидия трахоматис (Chlamydia trachomatis), микоплазма генитальная (Mycoplasma genitalium), трихомонада вагинальная (Trichomonas vaginalis), гонококк (Neisseria gonorrhoeae). Обследование проводится вне зависимости от наличия у женщины клинических и лабораторных признаков воспаления.
- Диагностика условно-патогенных микоплазм: качественное и количественное определение уреаплазма парвум (Ureaplasma parvum), уреаплазма уреалитикум (Ureaplasma urealyticum), микоплазма гоминис (Mycoplasma hominis). Данные микроорганизмы нередко являются причиной воспалительных заболеваний мочеполовой сферы. В зависимости от концентрации этих микроорганизмов и клинической картины, врач принимает решение о назначении терапии.
- Диагностика бактериального вагиноза: качественное и количественное определение общей бактериальной массы Bacteria, лактобактерий Lactobacillus spp., гарднерелла вагинальная (Gardnerella vaginalis) и атопобиум вагинальный (Atopobium vaginae). В зависимости от обнаруженного маркера бактериального вагиноза определяется дальнейшая тактика лечения.
- Диагностика аэробного вагинита: качественное и количественное определение Энтеробактерий (Enterobacteriaceae), стафилококков (Staphylococcus spp. ), стрептококков (Streptococcus spp.). В случаях, когда количество лактобактерий значительно снижено, а данные микроорганизмы составляют основную долю в бактериальной массе, мы говорим о преобладании аэробной флоры. Что позволяет правильно подобрать необходимые лекарственные препараты.
- Диагностика кандидозного вульвовагинита: качественное и количественное определение грибов рода Кандида: Candida albicans, Candida glabrata, Candida krusei, Candida parapsilosis/tropicalis. Расширение диагностического диапазона грибов рода Candida и их концентрация позволяет корректировать медикаментозное лечение в зависимости от обнаруженного вида.
 Обследование проводится вне зависимости от наличия у женщины клинических и лабораторных признаков воспаления.
Обследование проводится вне зависимости от наличия у женщины клинических и лабораторных признаков воспаления. ), стрептококков (Streptococcus spp.). В случаях, когда количество лактобактерий значительно снижено, а данные микроорганизмы составляют основную долю в бактериальной массе, мы говорим о преобладании аэробной флоры. Что позволяет правильно подобрать необходимые лекарственные препараты.
), стрептококков (Streptococcus spp.). В случаях, когда количество лактобактерий значительно снижено, а данные микроорганизмы составляют основную долю в бактериальной массе, мы говорим о преобладании аэробной флоры. Что позволяет правильно подобрать необходимые лекарственные препараты.Симптомы и лечение гарднереллеза у мужчин в Санкт-Петербурге
Автор статьи: КМН, врач уролог-андролог, специалист УЗД, зав. отделением урологииРыльчиков И. В. Стаж работы: 21 год
Опубликовано: 05-08-2020
Обновлено: 22-06-2020
Гарднереллёз — воспалительное заболевание уретры и наружных половых органов у мужчин и влагалища у женщин, при котором в указанных органах растёт количество бактерий рода Гарднерелла.
Заболевание или норма?
Гарднереллёз (другое название вагинальный дисбактериоз) — заболевание больше присущее женскому организму, поскольку его причиной служит бактериальный фон влагалища, где гарднереллы должны содержаться в норме. Только дисбаланс этих бактерий приводит к гарднереллёзу у женщин. У мужчин это происходит гораздо реже и является лишь следствием заражения. В норме возбудитель гарднерёллеза выводится из мочеполового тракта мужчины на протяжении двух-трех суток после проникновения. Сниженный иммунитет, переохлаждения, сопутствующие заболевания способствуют росту количества гарднерелл и приводит к воспалительному процессу в уретре.
Цены на услуги
Первичный приём уролога ДЛЯ МУЖЧИН + УЗИ предстательной железы (оценка жалоб, сбор анамнеза, при необходимости ректальное пальцевое исследование, узи предстательной железы у мужчин)
Первичный прием – обращение к врачу конкретной специальности в первый раз. Включает беседу с пациентом, первичный осмотр, сбор анамнеза, при необходимости ректальное пальцевое исследование, узи предстательной железы.
Цена действительна с 01.02.2021
Цена не действительна на прием в филиалах у м. пр. Большевиков и у м. пр. Просвещения
Включает беседу с пациентом, первичный осмотр, сбор анамнеза, при необходимости ректальное пальцевое исследование, узи предстательной железы.
Цена действительна с 01.02.2021
Цена не действительна на прием в филиалах у м. пр. Большевиков и у м. пр. Просвещения
Первичный приём зав. отделением урологии Рыльчиковым И.В. + УЗИ (включает беседу с пациентом, первичный осмотр, сбор анамнеза, при необходимости ректальное пальцевое исследование, узи предстательной железы)
Первичный прием – обращение к врачу конкретной специальности в первый раз.
Записаться на приемЗаражение
Поскольку гарднереллы являются частью нормальной микрофлоры женщины, то проникнуть в организм мужчины они могут только во время незащищённого секса. Бактерии попадают на слизистую оболочку уретры, могут подняться вверх по половым путям и под воздействием предрасполагающих факторов вызвать вялотекущий воспалительный процесс. Инфицированный мужчина в свою очередь может стать источником заражения для другой партнёрши.
Бактерии попадают на слизистую оболочку уретры, могут подняться вверх по половым путям и под воздействием предрасполагающих факторов вызвать вялотекущий воспалительный процесс. Инфицированный мужчина в свою очередь может стать источником заражения для другой партнёрши.
ВАЖНО! У женщины гарднереллёз не может возникнуть, как говориться, на пустом месте — он тесно связан с факторами риска венерических болезней. Поэтому женщинам с гарднереллёзом необходимо обследоваться на основные заболевания, передаваемые половым путём.
Симптомы
У мужчин симптомы гарднереллёза, как правило, бывают не яркими. У подавляющего большинства представителей сильного пола гарднерелла не вызывает никаких внешних изменений. Но иногда гарднереллёз является причиной развития уретрита — воспаления мочеиспускательного канала. В этом случае в выделениях из полового члена ощущается специфический «рыбный» запах, возможны небольшие выделения беловато-серого цвета. Кроме того, могут возникать жжение, зуд, неприятные ощущения при мочеиспускании.
ФАКТ: вне зависимости от наличия или отсутствия симптомов гарднереллёза заражённый мужчина всё равно становится источником инфицирования своей партнерши.
Осложнения
Если лечение гарднереллёза не было вовремя проведено, есть вероятность развития осложнений. Среди таких последствий гарднереллёза у мужчин — уретрит, крайне редко — простатит, эпидидимит, и др.
У женщин гарднереллёз считают фактором риска воспаления наружных половых органов, влагалища и шейки матки. Это проявляется болью, резями, жжением в наружных половых органах, сухостью во влагалище и болью во время полового акта. Большое количество гарднерелл снижает иммунитет всех органов малого таза, что способствует развитию более серьёзных и трудноизлечимых заболеваний. Мы говорим здесь о женских осложнениях как о факторе ответственности мужчины за свою партнёршу и будущее потомство.
Записаться на приемДиагностика
Диагностика гарднереллёза не представляет для специалиста особых трудностей. Диагноз ставится на основании исследования мазка из уретры мужчин. Надо заметить, что желательно помимо анализа на гарднереллёз, сдать материал и на скрытые инфекции, чтобы исключить или подтвердить их наличие и в зависимости от результата назначать лечение.
Диагноз ставится на основании исследования мазка из уретры мужчин. Надо заметить, что желательно помимо анализа на гарднереллёз, сдать материал и на скрытые инфекции, чтобы исключить или подтвердить их наличие и в зависимости от результата назначать лечение.
ВАЖНО! Анализы нужно сдавать обоим — и мужчине, и женщине, особенно если они сменили одного или нескольких половых партнёров.
Лечение
Лечение гарднереллёза включает применение антибиотиков и составляет в среднем 1-3 недели. На этот период необходимо отказаться от половых связей и соблюдать элементарную диету, исключающую острую, жареную, копченую, пряную пищу и алкоголь. После проведённой терапии проводятся контрольные исследования для исключения рецидивов заболевания. Обязательным условием является проведение лечения гарднереллёза всех половых партнёров, иначе оно не будет эффективным.
ФАКТ: даже успешно проведённая терапия гарднереллёза не гарантирует отсутствие рецидивов у женщины, поскольку заболевание связано с общим состоянием здоровья, силой иммунитета и может быть спровоцировано такими факторами, как приём гормональных препаратов, стрессы, сопутствующие инфекции и неправильное питание.
Профилактика.
Профилактика гарднереллеза у мужчин состоит, прежде всего, в надёжном предохранении при новых половых контактах, заботе о своём иммунитете и непременном лечении постоянной партнёрши.
Другие статьи автора
Врачи
Все врачи| ПЦР-ДИАГНОСТИКА (мазок, соскоб из урогенитального тракта) | |||
| Бактериальный вагиноз (Gardnerella vaginalis/ Lactobacillus sp/ Atopobium vaginae/ количество клеток), определение ДНК, кол.ан. | до 5 д. | 1090 р. | |
| Вирус папилломы человека высокого канцерогенного риска (HPV 16, 18, 31, 33, 35, 39, 45, 51, 52,56, 58, 59), качественное определение ДНК с указанием типа вируса (генотипирование) | до 9 д. |
820 р. | |
| Вирус папилломы человека высокого канцерогенного риска, без определения типа (HPV 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68), определение ДНК, кач.ан. | до 5 д. | 540 р. | |
| Вирус папилломы человека высокого канцерогенного риска, без определения типа (у женщин) (HPV 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68), определение ДНК, кол.ан. | до 7 д. | 1210 р. | |
| Вирус папилломы человека, типы 16,18 (HPV 16,18), определение ДНК, кач.ан. | до 5 д. | 330 р. | |
| Вирус папилломы человека, типы 16,18 (HPV 16,18), определение ДНК, кол.ан. | до 7 д. |
610 р. | |
| Вирус папилломы человека, типы 6,11 (HPV 6,11), определение ДНК, кач.ан. | до 5 д. | 330 р. | |
| Вирус простого герпеса 1 и 2 типа (HSV 1,2), определение ДНК, кач.ан. | до 4 д. | 330 р. | |
| Возбудители кандидоза (Candida albicans/Candida glabrata/Candida krusei/Candida parapsilosis /Candida tropicalis), определение ДНК, кол.ан. | до 5 д. | 600 р. | |
| Гарднерелла вагиналис (Gardnerella vaginalis), определение ДНК, кач.ан. | до 4 д. | 330 р. | |
Кандида Альбиканс (Candida albicans), определение ДНК, кач. ан. ан. |
до 4 д. | 330 р. | |
| Микоплазма гениталиум (Mycoplasma genitalium), определение ДНК, кач.ан. | до 4 д. | 330 р. | |
| Микоплазма гениталиум (Mycoplasma genitalium), определение ДНК, кол.ан. | до 5 д. | 720 р. | |
| Микоплазма хоминис (Mycoplasma hominis), определение ДНК, кач.ан. | до 4 д. | 330 р. | |
| Микоплазма хоминис (Mycoplasma hominis), определение ДНК, кол.ан. | до 5 д. | 680 р. | |
Нейссерия гонореи (Neisseria gonorrhoeae), определение ДНК, кач. ан. ан. |
до 4 д. | 330 р. | |
| Нейссерия гонореи (Neisseria gonorrhoeae), определение ДНК, кол.ан. | до 5 д. | 720 р. | |
| Типирование уреаплазмы (U.urealyticum / U. parvum), определение ДНК, кач.ан. | до 4 д. | 330 р. | |
| Типирование уреаплазмы (U.urealyticum / U. parvum), определение ДНК, кол.ан. | до 5 д. | 550 р. | |
| Трепонема паллидум (Treponema pallidum)(Сифилис), определение ДНК, кач.ан. | до 4 д. | 330 р. | |
Трихомонас вагиналис (Trichomonas vaginalis), определение ДНК, кач. ан. ан. |
до 4 д. | 330 р. | |
| Трихомонас вагиналис (Trichomonas vaginalis), определение ДНК, кол.ан. | до 5 д. | 720 р. | |
| Урогенитальные инфекции (ПЦР 12 соскоб) (Хламидия трахоматис, Микоплазма гениталиум, Трихомонас вагиналис, Нейссерия гонорея, Микоплазма хоминис, Уреаплазма уреалитикум, Уреаплазма парвум, Гарднерелла вагиналис, Кандида альбиканс, Цитомегаловирус, Вирус простого герпеса 1 и 2), кач.ан. | до 4 д. | 2360 р. | |
Урогенитальные инфекции (ПЦР 15 соскоб) (Хламидия трахоматис, Микоплазма гениталиум, Трихомонас вагиналис, Нейссерия гонорея, Микоплазма хоминис, Уреаплазма уреалитикум, Уреаплазма парвум, Гарднерелла вагиналис, Кандида альбиканс, Цитомегаловирус, Вирус простого герпеса 1 и 2, Трепонема паллидум, Вирус папилломы человека 6/11), кач. ан. ан. |
до 4 д. | 2860 р. | |
| Урогенитальные инфекции (ПЦР 7 соскоб) (Хламидия трахоматис, Микоплазма гениталиум, Трихомонас вагиналис, Нейссерия гонорея, Микоплазма хоминис, Уреаплазма уреалитикум, Уреаплазма парвум), кач.ан. | до 4 д. | 1450 р. | |
| Урогенитальные инфекции (ПЦР 9 соскоб) (Хламидия трахоматис, Микоплазма гениталиум, Трихомонас вагиналис, Нейссерия гонорея, Микоплазма хоминис, уреаплазма уреалитикум, Уреаплазма парвум, Гарднерелла вагиналис, Кандида альбиканс), кач.ан. | до 4 д. | 1920 р. | |
Урогенитальные инфекции у женщин (Нейссерия гонорея, Хламидия трахоматис, Микоплазма гениталиум, Трихомонас вагиналис, Уреаплазма уреалитикум, Уреаплазма парвум, Микоплазма хоминис, Кандида альбиканс/глабрата/крузей/парапсилозис и тропикалис, Бактериальный вагиноз) определение ДНК, кол. ан. (соскоб) ан. (соскоб) |
до 5 д. | 2700 р. | |
| Урогенитальные инфекции у мужчин (Нейссерия гонорея, Хламидия трахоматис, Микоплазма гениталиум, Трихомонас вагиналис, Уреаплазма уреалитикум, Уреаплазма парвум, Микоплазма хоминис, Кандида альбиканс/глабрата/крузей/парапсилозис и тропикалис), кол.ан.(соскоб) | до 5 д. | 2400 р. | |
| Фемофлор 16 (Исследование биоценоза урогенитального тракта) | до 5 д. | 2200 р. | |
| Фемофлор 8 (Исследование биоценоза урогенитального тракта) | до 5 д. | 1100 р. | |
| Фемофлор скрин (Исследование биоценоза урогенитального тракта) | до 5 д. |
1650 р. | |
| Флороценоз и NCMT (ДНК Candida albicans, ДНК Candida glabrata, ДНК Candida krusei, ДНК Candida parapsilosis/tropicalis, ДНК Ureaplasma parvum, ДНК Ureaplasma urealyticum, ДНК Mycoplasma hominis, ДНК Cardnerella vaginalis, ДНК Atopobium vaginae, ДНК Enterobacteriaceae, ДНК Staphylococcus spp., ДНК Streptococcus spp., ДНК Lactobacillus spp., ДНК Bacteria spp., ДНК Neisseria gonorrhoeae, ДНК Chlamydia trachomatis, ДНК Mycoplasma genitalium, ДНК Trichomonas vaginalis), кол.ан. | до 3 д. | 2170 р. | |
| Хламидия трахоматис (Chlamydia trachomatis), определение ДНК, кач.ан. | до 4 д. | 330 р. | |
| Хламидия трахоматис (Chlamydia trachomatis), определение ДНК, кол.ан. | до 5 д. |
720 р. | |
| Цитомегаловирус (Cytomegalovirus), определение ДНК, кач.ан. | до 4 д. | 330 р. | |
Бактериальная ДНК — роль плазмид — Science Learning Hub
Как и другие организмы, бактерии используют двухцепочечную ДНК в качестве своего генетического материала. Однако бактерии организуют свою ДНК иначе, чем более сложные организмы.
Бактериальная ДНК — круговая хромосома плюс плазмиды
ДНК большинства бактерий содержится в одной кольцевой молекуле, называемой бактериальной хромосомой. Хромосома вместе с несколькими белками и молекулами РНК образует структуру неправильной формы, называемую нуклеоидом.Он находится в цитоплазме бактериальной клетки.
Помимо хромосомы, бактерии часто содержат плазмиды — маленькие кольцевые молекулы ДНК. Бактерии могут улавливать новые плазмиды из других бактериальных клеток (во время конъюгации) или из окружающей среды. Они также могут легко потерять их — например, когда бактерия делится на две части, одна из дочерних клеток может не получить плазмиду.
Они также могут легко потерять их — например, когда бактерия делится на две части, одна из дочерних клеток может не получить плазмиду.
Каждая плазмида имеет свою собственную «точку начала репликации» — участок ДНК, который обеспечивает ее репликацию (копирование) бактерией-хозяином.По этой причине плазмиды могут копировать себя независимо от бактериальной хромосомы, поэтому в одной бактериальной клетке может быть много копий плазмиды — даже сотни.
Плазмиды помогают бактериям пережить стресс
Плазмиды содержат всего несколько генов, но они имеют большое значение для бактерии-хозяина. Гены обычно не важны для повседневного выживания бактерии — вместо этого они помогают бактерии преодолевать периодические стрессовые ситуации. Например, многие плазмиды содержат гены, которые при экспрессии делают бактерию-хозяин устойчивой к антибиотику (чтобы она не погибла при обработке этим антибиотиком).Другие плазмиды содержат гены, которые помогают хозяину переваривать необычные вещества или убивать другие типы бактерий.
Плазмиды сами по себе незаменимы
Сохранение плазмиды — тяжелая работа для бактериальной клетки, потому что для репликации ДНК (включая плазмидную ДНК) расходуется энергия. Однако, защищая своего бактериального хозяина от смерти, связанной со стрессом, плазмида максимизирует свои шансы на то, чтобы ее сохранить. В стрессовых условиях бактерии с плазмидой будут жить дольше — и у них будет больше возможностей передать плазмиду дочерним клеткам или другим бактериям.Бактерии без плазмиды с меньшей вероятностью выживут и размножаются.
Некоторые плазмиды принимают крайние меры, чтобы гарантировать, что они удерживаются в бактериях. Например, некоторые несут ген, производящий долгоживущий яд, и второй ген, создающий недолговечное противоядие. Эти плазмиды эффективно удерживают свою бактериальную клетку-хозяин в заложниках — если они когда-либо будут потеряны из клетки, они не смогут предоставить противоядие, и клетка умрет.
Плазмиды в биотехнологии — доставка ДНК
Плазмиды сыграли ключевую роль в развитии молекулярной биотехнологии. Они действуют как средства доставки или векторы для введения чужеродной ДНК в бактерии. Использование плазмид для доставки ДНК началось в 1970-х годах, когда ДНК других организмов была сначала «вырезана и вставлена» в определенные участки внутри плазмидной ДНК. Затем модифицированные плазмиды повторно вводили в бактерии.
Они действуют как средства доставки или векторы для введения чужеродной ДНК в бактерии. Использование плазмид для доставки ДНК началось в 1970-х годах, когда ДНК других организмов была сначала «вырезана и вставлена» в определенные участки внутри плазмидной ДНК. Затем модифицированные плазмиды повторно вводили в бактерии.
Почему плазмиды являются отличными векторами доставки ДНК
Спустя десятилетия после их первого использования плазмиды по-прежнему являются важнейшими лабораторными инструментами в биотехнологии:
- Ученые могут заставить бактерии удерживать их. Практически все плазмиды, которые используются для доставки ДНК, содержат гены устойчивости к антибиотикам. После обработки бактерий плазмидой ученые выращивают их в присутствии антибиотика. Только те клетки, которые содержат плазмиду, выживут, будут расти и воспроизводиться. Остальные будут убиты антибиотиком.
- Копируются самостоятельно. Плазмиды можно копировать множество раз, независимо от того, реплицирует ли бактериальный хозяин свою собственную ДНК, и каждый раз, когда реплицируется плазмидный вектор, то же самое происходит и с введенной ДНК, которую он содержит.
- Они круглые. ДНК , имеющая кольцевую форму, хорошо подходит для включения дополнительных последовательностей ДНК. Это потому, что его можно разрезать, не развалившись, а затем снова соединиться, как только будет встроена новая ДНК.

Узнайте больше о том, как добавить чужеродную ДНК к бактериям.
Бактериальная ДНК — обзор
7.19.2 Механизм антибактериального действия
Фторхинолоны нарушают синтез бактериальной ДНК, что приводит к быстрой гибели бактериальных клеток.Специфические мишени, ДНК-гираза и топоизомераза IV, являются членами класса ферментов топоизомеразы. 23 Обнаруженные спустя много времени после клинического использования фторхинолонов мишенями противомикробного действия являются ДНК-связывающие ферменты, которые контролируют топологию ДНК, необходимую для роста и деления клеток. Поскольку ДНК-гираза и топоизомераза IV являются внутриклеточными ферментами, хинолоны должны проникать через бактериальную мембрану и накапливаться в бактериальной клетке до концентрации, достаточной для обеспечения антибактериальной активности. Простая диффузия через бактериальную мембрану приводит к антимикробной активности у E. coli и S. aureus . 24 Однако у других грамотрицательных видов накопление хинолона и антибактериальное действие были связаны с активным поглощением поринами внешней мембраны. 25 Защитные механизмы, такие как низкомолекулярные насосы, также могут играть роль в снижении накопления хинолонов. Защитные системы могут резко снизить эффективность хинолонов у некоторых организмов, что приведет к устойчивости, что будет обсуждаться в следующем разделе.
Простая диффузия через бактериальную мембрану приводит к антимикробной активности у E. coli и S. aureus . 24 Однако у других грамотрицательных видов накопление хинолона и антибактериальное действие были связаны с активным поглощением поринами внешней мембраны. 25 Защитные механизмы, такие как низкомолекулярные насосы, также могут играть роль в снижении накопления хинолонов. Защитные системы могут резко снизить эффективность хинолонов у некоторых организмов, что приведет к устойчивости, что будет обсуждаться в следующем разделе.
В этом классе есть две группы топоизомераз, которые имеют различные биохимические механизмы и физиологические роли, хотя обе группы регулируют сверхспиральное состояние бактериальной ДНК. Природа или степень суперсильности контролирует начало репликации ДНК и, как следствие, транскрипцию многих бактериальных генов. Топоизомеразы типа I, которые включают топоизомеразу I и топоизомеразу III, не ингибируются хинолонами или другими антибиотиками. Топоизомеразы типа I катализируют расщепление одиночных цепей ДНК, вызывая разрывы, пассажи одиночных цепей и лигирование для снятия сверхспиральных скручиваний и декататированных взаимосвязанных кругов ДНК. 26 Топоизомеразы типа I устраняют отрицательные сверхспиральные изгибы ДНК, действие, которое противодействует активности ДНК-гиразы. Антимикробная активность фторхинолонов проявляется в основном за счет ингибирования ДНК-гиразы и топоизомеразы IV, которые являются топоизомеразами II типа. Топоизомеразы типа II катализируют расщепление, пассаж и повторное лигирование двухцепочечной ДНК.
Топоизомеразы типа I катализируют расщепление одиночных цепей ДНК, вызывая разрывы, пассажи одиночных цепей и лигирование для снятия сверхспиральных скручиваний и декататированных взаимосвязанных кругов ДНК. 26 Топоизомеразы типа I устраняют отрицательные сверхспиральные изгибы ДНК, действие, которое противодействует активности ДНК-гиразы. Антимикробная активность фторхинолонов проявляется в основном за счет ингибирования ДНК-гиразы и топоизомеразы IV, которые являются топоизомеразами II типа. Топоизомеразы типа II катализируют расщепление, пассаж и повторное лигирование двухцепочечной ДНК.
ДНК-гираза, которая была первой идентифицированной мишенью хинолонов, является важным (необходимым для жизни) ферментом бактерий. 27 Бактериальная ДНК обычно находится в равновесии между замкнутым кольцевым двухцепочечным состоянием (расслабленным) и сильно отрицательно свернутым состоянием (рис. 4). ДНК-гираза контролирует топологию бактериальной ДНК и функцию хромосом, поддерживая отрицательную суперспирализацию ДНК (сверхспиральные скручивания). Решающее значение для репликации ДНК, ДНК-гираза отвечает за ослабление положительного сверхспирального скручивания, которое накапливается перед активной вилкой репликации ДНК. 28 Кроме того, ДНК-гираза участвует в изгибании и складывании ДНК и удалении узлов (рис. 4).Активный фермент состоит из четырех субъединиц, двух субъединиц А и двух субъединиц В, которые являются продуктами генов gyrA и gyrB соответственно. 29 Субъединица A является сайтом связывания хинолона и требует активного сайта Tyr-122 для каталитической активности. Рентгеновские структуры были получены для: (1) фрагмента N-концевого домена (доменов «разрыв-соединение» и области, определяющей устойчивость к хинолонам (QRDR)) субъединицы A 30 ; и (2) дрожжевая топоизомераза II. 31 АТФ-связывающий домен находится в субъединице В, которая опосредует АТФазную активность ДНК-гиразы.Кристаллическая структура доступна для N-концевого домена 43 кДа (АТФаза и домены захвата ДНК) субъединицы B.
Решающее значение для репликации ДНК, ДНК-гираза отвечает за ослабление положительного сверхспирального скручивания, которое накапливается перед активной вилкой репликации ДНК. 28 Кроме того, ДНК-гираза участвует в изгибании и складывании ДНК и удалении узлов (рис. 4).Активный фермент состоит из четырех субъединиц, двух субъединиц А и двух субъединиц В, которые являются продуктами генов gyrA и gyrB соответственно. 29 Субъединица A является сайтом связывания хинолона и требует активного сайта Tyr-122 для каталитической активности. Рентгеновские структуры были получены для: (1) фрагмента N-концевого домена (доменов «разрыв-соединение» и области, определяющей устойчивость к хинолонам (QRDR)) субъединицы A 30 ; и (2) дрожжевая топоизомераза II. 31 АТФ-связывающий домен находится в субъединице В, которая опосредует АТФазную активность ДНК-гиразы.Кристаллическая структура доступна для N-концевого домена 43 кДа (АТФаза и домены захвата ДНК) субъединицы B. 32
32
Рис. 4. Суперспирализация ДНК и образование узлов.
На основании как биохимических, так и структурных данных каталитический цикл ДНК-гиразы можно выразить как открытие и закрытие серии «молекулярных ворот», связанных с гидролизом АТФ. 33 ДНК-гираза вступает в контакт и оборачивает сегмент ДНК длиной 120 п.н. в положительную суперспираль, в результате чего образуется нековалентный комплекс связывания фермент-ДНК (рис. 5).Следующим этапом является образование расщепляемого комплекса, в котором Tyr-122 из субъединиц A ковалентно связывается с ДНК через фосфодиэфирное расщепление в типичной шахматной конфигурации из 4 оснований. На этом этапе возможен проход прядей. Переход цепи происходит, когда связывание АТФ вызывает структурные изменения с сопутствующим транспортом ДНК через временно расщепленную ДНК, связанную в ферментном комплексе. Наконец, концы ДНК повторно соединяются (повторно запечатываются), что приводит к образованию отрицательной суперспирали. Гидролиз АТФ запускает регенерацию конформации предплечья, а диссоциация АДФ регенерирует активный фермент, завершая ферментативный цикл.
Гидролиз АТФ запускает регенерацию конформации предплечья, а диссоциация АДФ регенерирует активный фермент, завершая ферментативный цикл.
Рис. 5. ДНК-гираза: образование расщепляемого комплекса, пассаж цепи и связывание хинолона.
Вторая мишень фторхинолонов, топоизомераза IV, отвечает за разделение связанных (связанных) кольцевых дочерних молекул ДНК, которые являются продуктом репликации ДНК. 34 Как только две вновь реплицированные молекулы ДНК разделены в процессе расщепления, пассажа и повторного лигирования, бактериальная клетка приготавливается для деления с выделением одной копии ДНК в каждой из вновь образованных дочерних клеток.Таким образом, хотя каталитическая активность ДНК-гиразы и топоизомеразы IV механически схожа, они играют разные и важные роли в репликации ДНК и пролиферации клеток. 35 Топоизомераза IV, как и ДНК-гираза, представляет собой тетрамер, состоящий из двух пар из двух субъединиц, кодируемых генами parC и parE . Будучи тесно связанными с ДНК-гиразой по структуре и функциям, гены и продукты демонстрируют близкую гомологию; parC для gyrA и parE для gyrB . 36 Основное различие между топоизомеразой IV и ДНК-гиразой заключается в том, как ДНК связывается с ферментами. В ДНК-гиразе нековалентные связывающие взаимодействия между ДНК и ферментом поддерживают тесную ассоциацию или «обертывание» ДНК вокруг белка. «Обертывание» способствует «внутримолекулярному» прохождению цепи. Альтернативно, топоизомераза IV не проявляет «оборачивания» ДНК и, таким образом, способствует «межмолекулярному» пассажу цепи. Делеция части С-концевого домена («оборачивающая» часть) GyrA превращает ДНК-гиразу в фермент, который не может катализировать суперспирализацию, но обладает сильной активностью декатенации, подобной топоизомеразе IV. 37
Будучи тесно связанными с ДНК-гиразой по структуре и функциям, гены и продукты демонстрируют близкую гомологию; parC для gyrA и parE для gyrB . 36 Основное различие между топоизомеразой IV и ДНК-гиразой заключается в том, как ДНК связывается с ферментами. В ДНК-гиразе нековалентные связывающие взаимодействия между ДНК и ферментом поддерживают тесную ассоциацию или «обертывание» ДНК вокруг белка. «Обертывание» способствует «внутримолекулярному» прохождению цепи. Альтернативно, топоизомераза IV не проявляет «оборачивания» ДНК и, таким образом, способствует «межмолекулярному» пассажу цепи. Делеция части С-концевого домена («оборачивающая» часть) GyrA превращает ДНК-гиразу в фермент, который не может катализировать суперспирализацию, но обладает сильной активностью декатенации, подобной топоизомеразе IV. 37
Многочисленные исследования изучали молекулярную целевую специфичность хинолонов в широком спектре бактерий. У грамотрицательных видов, таких как E. coli , первичной мишенью, по-видимому, является ДНК-гираза. 38 Данные свидетельствуют о том, что хинолоны связываются с комплексом фермент-ДНК после расщепления цепи, таким образом стабилизируя комплекс и ингибируя стадию лигирования цепи. Стабилизированный комплекс создает разрыв ДНК, который клетка плохо приспособлена для восстановления.Предполагается, что стабилизированный или «захваченный» комплекс действует как клеточный яд, что может помочь объяснить несоответствие между эффективностью хинолона на ферментативном и клеточном уровнях (часто МИК намного ниже, чем значения IC 50 или CC 50 ). 39 Образование стабилизированного тройного комплекса приводит к ингибированию синтеза ДНК и роста клеток за счет блокировки репликационной вилки. Затем происходит летальное разрушение ДНК (двухцепочечные разрывы), что требует синтеза РНК и белка для восстановления повреждений.В результате хинолоны как класс обладают бактерицидным действием. В ответ на клеточное повреждение бактерии запускают путь SOS; экспрессия набора генов, которые включают репарацию ДНК, рекомбинацию ДНК (например, нежизнеспособные клетки) и мутагенез (например, потенциальную устойчивость).
coli , первичной мишенью, по-видимому, является ДНК-гираза. 38 Данные свидетельствуют о том, что хинолоны связываются с комплексом фермент-ДНК после расщепления цепи, таким образом стабилизируя комплекс и ингибируя стадию лигирования цепи. Стабилизированный комплекс создает разрыв ДНК, который клетка плохо приспособлена для восстановления.Предполагается, что стабилизированный или «захваченный» комплекс действует как клеточный яд, что может помочь объяснить несоответствие между эффективностью хинолона на ферментативном и клеточном уровнях (часто МИК намного ниже, чем значения IC 50 или CC 50 ). 39 Образование стабилизированного тройного комплекса приводит к ингибированию синтеза ДНК и роста клеток за счет блокировки репликационной вилки. Затем происходит летальное разрушение ДНК (двухцепочечные разрывы), что требует синтеза РНК и белка для восстановления повреждений.В результате хинолоны как класс обладают бактерицидным действием. В ответ на клеточное повреждение бактерии запускают путь SOS; экспрессия набора генов, которые включают репарацию ДНК, рекомбинацию ДНК (например, нежизнеспособные клетки) и мутагенез (например, потенциальную устойчивость). Однако в некоторых случаях ингибирование синтеза ДНК может быть не связано с бактерицидной активностью, что открывает возможность альтернативных путей или дополнительных факторов, участвующих в механизме действия. 40
Однако в некоторых случаях ингибирование синтеза ДНК может быть не связано с бактерицидной активностью, что открывает возможность альтернативных путей или дополнительных факторов, участвующих в механизме действия. 40
У грамотрицательных видов, таких как E.coli , обработка хинолоном генерирует устойчивые штаммы, которые имеют специфические одиночные мутации в gyrA и gyrB . Эти данные показывают, что стабилизация ДНК-гиразы является основным механизмом действия в E. coli . Эти мутации придают устойчивость за счет снижения сродства связывания ДНК-гиразы с хинолоном. 41 В отличие от грамотрицательных бактерий, устойчивость к хинолонам у грамположительных, таких как S. pneumoniae и S. aureus , как правило, является результатом единичных мутаций в генах топоизомеразы IV, parC и parE. в S.pneumoniae , что позволяет предположить, что топоизомераза IV является основной хинолоновой мишенью у грамположительных видов. 38,42 При исследовании ингибирования ферментативного катализа батареей хинолонов в E. coli и S. aureus наиболее чувствительная топоизомераза у каждого вида следовала ожидаемой тенденции: в E. coli ДНК Гираза имела преимущество перед топоизомеразой IV в 15-27 раз, в то время как в S. aureus топоизомераза IV имела преимущество в 2-21 раз. 43 Однако структура хинолона играет роль в различении мишеней топоизомеразы. Недавние исследования новых хинолонов продемонстрировали, что субъединица gyrA ДНК-гиразы является основной мишенью для моксифлоксацина и гатифлоксацина в S. pneumoniae . 44 Основываясь на этих наблюдениях, изменения в замещении в позициях хинолона C-7 или C-8 могут играть значительную роль в целевом предпочтении. Хотя существует общая тенденция к специфичности мишеней между грамположительными и грамотрицательными видами, первичной мишенью топоизомеразы для данной бактерии является фермент, наиболее чувствительный к конкретному хинолону.
38,42 При исследовании ингибирования ферментативного катализа батареей хинолонов в E. coli и S. aureus наиболее чувствительная топоизомераза у каждого вида следовала ожидаемой тенденции: в E. coli ДНК Гираза имела преимущество перед топоизомеразой IV в 15-27 раз, в то время как в S. aureus топоизомераза IV имела преимущество в 2-21 раз. 43 Однако структура хинолона играет роль в различении мишеней топоизомеразы. Недавние исследования новых хинолонов продемонстрировали, что субъединица gyrA ДНК-гиразы является основной мишенью для моксифлоксацина и гатифлоксацина в S. pneumoniae . 44 Основываясь на этих наблюдениях, изменения в замещении в позициях хинолона C-7 или C-8 могут играть значительную роль в целевом предпочтении. Хотя существует общая тенденция к специфичности мишеней между грамположительными и грамотрицательными видами, первичной мишенью топоизомеразы для данной бактерии является фермент, наиболее чувствительный к конкретному хинолону.
Бактериальные геномы | Кафедра микробиологии
Все живые организмы содержат ДНК. Эта удивительная макромолекула кодирует всю информацию, необходимую для программирования деятельности клетки, включая воспроизводство, метаболизм и другие специализированные функции. ДНК состоит из двух цепей дезоксинуклеотидов. Каждый дезоксинуклеотид содержит фосфат, 5-углеродный сахар (2-дезоксирибозу) и одно из четырех азотистых оснований: аденин, цитозин, тимин или гуанин. Фосфат и сахар составляют основу каждой цепи ДНК, а основания отвечают за удерживание двух цепей вместе посредством водородных связей в структуре, называемой двойной спиралью (см. Рисунок).Порядок оснований в цепи ДНК содержит закодированную генетическую информацию. Вся ДНК, обнаруженная в организме, вместе называется геномом. Геном человека состоит из 23 пар линейных хромосом и примерно 3000 мегабаз (МБ) ДНК, в то время как геном бактерии Escherichia coli состоит из одной кольцевой хромосомы размером 4,6 МП. Изучая геномы бактерий, мы можем лучше понять их метаболические возможности, их способность вызывать болезни, а также их способность выживать в экстремальных условиях.Многие из хорошо изученных бактериальных модельных организмов, такие как E. coli , имеют единственную кольцевую хромосому. Однако достижения в области молекулярной генетики показали, что бактерии обладают более сложным расположением своего генетического материала, чем просто одна круговая хромосома на клетку. Некоторые бактериальные геномы состоят из нескольких хромосом и / или плазмид, и многие бактерии содержат несколько копий своего генома на клетку. Ниже приведены несколько примеров бактерий с необычным геномом.
Изучая геномы бактерий, мы можем лучше понять их метаболические возможности, их способность вызывать болезни, а также их способность выживать в экстремальных условиях.Многие из хорошо изученных бактериальных модельных организмов, такие как E. coli , имеют единственную кольцевую хромосому. Однако достижения в области молекулярной генетики показали, что бактерии обладают более сложным расположением своего генетического материала, чем просто одна круговая хромосома на клетку. Некоторые бактериальные геномы состоят из нескольких хромосом и / или плазмид, и многие бактерии содержат несколько копий своего генома на клетку. Ниже приведены несколько примеров бактерий с необычным геномом.
Терминология генома
Kb / Mb — килобаза (Kb) — это 1000 оснований ДНК, а мегабаза (Mb) — это 1000000 оснований.
Круглая хромосома · ДНК расположена по замкнутому кругу, который имеет отрицательную сверхспиральную спираль, что делает возможным компактную природу многих бактериальных геномов.
Линейная хромосома · Незамкнутая хромосома с инвертированными повторами на концах, похожая на телеомеры в хромосомах эукариот.
Плазмида · Внехромосомная ДНК, которая реплицируется независимо от хромосомы и регулирует собственную репликацию.
Мегаплазмида · Очень большая плазмида размером от 100 до 1700 килобайт.
Deinococcus radiodurans был впервые обнаружен в 1956 году Артуром В. Андерсоном. Осматривая испорченное мясо, он заметил, что образуются красноватые колонии, несмотря на то, что мясо было стерилизовано мегарадами радиации! Этот радиационно-устойчивый организм получил название Deinococcus radiodurans , что буквально означает «странная ягода, выдерживающая радиацию».« Deinococcus radiodurans способен выдерживать радиационное воздействие до 1 500 000 рад! Это в 3 000 раз больше, чем количество радиационного облучения, которое могло бы убить человека. Ионизирующее излучение вызывает двухцепочечные разрывы в ДНК. У клеток есть механизмы для их восстановления повреждения, но если сделано слишком много разрывов, сшивание ДНК в правильном порядке может подавить механизмы репарации ДНК клетки.Каким-то образом, D. radiodurans обладает способностью восстанавливать разрушенный геном.Геном D. radiodurans необычен тем, что он состоит из двух хромосом, мегаплазмиды и небольшой плазмиды. Кроме того, при делении клеток D. radiodurans и они не сразу полностью отделяются друг от друга, поэтому клетки часто существуют в виде тетрад (см., Например, фото). Хотя механизмы, с помощью которых D. radiodurans способен выдерживать высокие дозы радиации, все еще исследуются, предполагается, что благодаря наличию нескольких копий его генома и генетическому обмену между клетками в тетраде, D.radiodurans способен бороться с множественными разрывами ДНК, вызванными высокими уровнями радиации.
Ионизирующее излучение вызывает двухцепочечные разрывы в ДНК. У клеток есть механизмы для их восстановления повреждения, но если сделано слишком много разрывов, сшивание ДНК в правильном порядке может подавить механизмы репарации ДНК клетки.Каким-то образом, D. radiodurans обладает способностью восстанавливать разрушенный геном.Геном D. radiodurans необычен тем, что он состоит из двух хромосом, мегаплазмиды и небольшой плазмиды. Кроме того, при делении клеток D. radiodurans и они не сразу полностью отделяются друг от друга, поэтому клетки часто существуют в виде тетрад (см., Например, фото). Хотя механизмы, с помощью которых D. radiodurans способен выдерживать высокие дозы радиации, все еще исследуются, предполагается, что благодаря наличию нескольких копий его генома и генетическому обмену между клетками в тетраде, D.radiodurans способен бороться с множественными разрывами ДНК, вызванными высокими уровнями радиации.
Azotobacter vinelandii — большая почвенная облигатная аэробная бактерия, способная фиксировать азот. Кроме того, A. vinelandii может метаболизировать большое количество углеводов, органических кислот и спиртов. Количество геномов в отдельной клетке зависит от стадии роста клеток. Во время экспоненциального роста A.Клетки vinelandii обычно содержат от 2 до 4 копий своей хромосомы. Однако в стационарной фазе количество хромосом в отдельной клетке может увеличиваться до 50–100. Эта уникальная пластичность числа копий генома не совсем понятна, и необходимы дальнейшие исследования, чтобы лучше понять преимущества накопления большого количества хромосом в этих клетках во время стационарной фазы.
Buchnera spp.Эти бактерии являются внутриклеточными симбионтами некоторых видов тлей.Эта мутуалистическая связь между тлей и бактерией возникла миллионы лет назад. Хотя Buchnera тесно связана с E. coli , ее геном составляет примерно одну седьмую от размера генома E. coli . У одного вида Buchnera геном состоит из одной хромосомы размером 640 килобаз и двух плазмид, которые кодируют пути биосинтеза нескольких аминокислот. Было показано, что количество копий генома в клетках Buchnera связано со стадией развития тли-хозяина; по мере того, как тля вступает во взрослую жизнь, количество геномных копий в отдельных клетках Buchnera увеличивается.По мере старения тли-хозяина число копий генома у Buchnera уменьшается. Было высказано предположение, что это колебание числа копий может быть связано с тем, что бактерия очищает свои геномы от вредных мутаций, обеспечивая передачу только жизнеспособных хромосом следующему поколению тлей.
Agrobacterium tumefaciensЭти повсеместно распространенные грамотрицательные подвижные палочковидные почвенные бактерии являются возбудителем коронно-желчной болезни растений. Agrobacterium tumefaciens считается естественным генным инженером, поскольку она способна переносить ДНК от себя в клетки растений. Геном размером примерно 5,7 мегабаз (МБ) состоит из кольцевой хромосомы, линейной хромосомы и двух плазмид. Одна из плазмид, называемая плазмидой Ti для плазмиды, индуцирующей опухоль, отвечает за вирулентность A. tumefaciens .
Epulopiscium spp.Epulopiscium spp.являются кишечными симбионтами некоторых видов рыб-хирургов, принадлежащих к семейству Acanthuridae. Некоторые морфотипы Epulopiscium могут достигать длины более 0,5 мм! Это изображение представляет собой окрашенные DAPI клетки Epulopiscium . DAPI — это ДНК-специфическое пятно, и весь синий цвет, который вы видите в этих клетках, на самом деле ДНК. Анализы с использованием количественной ПЦР в реальном времени позволяют предположить, что Epulopiscium содержит десятки тысяч копий его генома. Такое количество копий беспрецедентно для бактерий и может отражать клеточную адаптацию, которая позволяет Epulopiscium поддерживать такой большой размер клеток.Имея тысячи копий своего генома, Epulopiscium может быть в состоянии синтезировать макромолекулы рядом с тем местом, где они необходимы в клетке, преодолевая ограничения, налагаемые коэффициентами диффузии малых молекул и биомолекул.
Список литературы
Интересующие сайты
Генетика — Медицинская микробиология — Книжная полка NCBI
Генетическая информация в микробах
Генетическим материалом бактерий и плазмид является ДНК. Бактериальные вирусы (бактериофаги или фаги) имеют ДНК или РНК в качестве генетического материала.Два основных Функции генетического материала — репликация и экспрессия. Генетический материал должен точно реплицироваться, чтобы потомство унаследовало все специфические генетические детерминанты (генотип) родительского организма. Выражение конкретных генетических материал в определенном наборе условий роста определяет наблюдаемые характеристика (фенотип) организма. Бактерии имеют мало структурных или особенности развития, которые можно легко наблюдать, но у них есть широкий спектр биохимические возможности и модели восприимчивости к противомикробным агентам или бактериофаги.Эти последние характеристики часто выбираются как наследуемые. черты, подлежащие анализу в исследованиях бактериальной генетики.
Структура нуклеиновой кислоты
Нуклеиновые кислоты — это большие полимеры, состоящие из повторяющихся нуклеотидных единиц (). Каждый нуклеотид содержит один фосфатная группа, один пентозный или дезоксипентозный сахар и один пурин или пиримидин основание. В ДНК сахар — это D-2-дезоксирибоза; в РНК сахар — это D-рибоза. В ДНК пуриновые основания представляют собой аденин (A) и гуанин (G), а пиримидиновые основания представляют собой тимин (Т) и цитозин (С).В РНК урацил (U) заменяет тимин. Химически модифицированные пуриновые и пиримидиновые основания обнаружены у некоторых бактерий и бактериофаги. Повторяющаяся структура полинуклеотидов включает чередование остатки сахара и фосфата, с фосфодиэфирными связями, связывающими 3′-гидроксильная группа одного нуклеотидного сахара к 5′-гидроксилу группа соседнего нуклеотидного сахара. Эти асимметричные фосфодиэфирные связи определяют полярность полинуклеотидной цепи. Основание пурина или пиримидина связан у 1′-углеродного атома каждого остатка сахара и выступает из повторяющийся сахарно-фосфатный остов.Двухцепочечная ДНК является спиральной, а две нити спирали антипараллельны. Двойная спираль стабилизирована водородные связи между пуриновыми и пиримидиновыми основаниями на противоположных цепях. В каждое положение, A на одной нити пары двумя водородными связями с T на противоположная нить, или G пары тремя водородными связями с C. Две нити поэтому двойные спирали ДНК комплементарны. Из-за взаимодополняемости двухцепочечная ДНК содержит эквимолярные количества пуринов (A + G) и пиримидины (T + C), где A равно T, а G равно C, но моль доля G + C в ДНК широко варьирует у разных бактерий.Информация в нуклеиновых кислотах кодируется упорядоченной последовательностью нуклеотидов. вдоль полинуклеотидной цепи, а в двухцепочечной ДНК последовательность каждого strand определяет, какой должна быть последовательность комплементарной цепи. В степень гомологии последовательностей между ДНК из разных микроорганизмов является самый строгий критерий для определения того, насколько тесно они связаны.
Рисунок 5-1
Двойная спиральная структура ДНК. На схеме представлена структура ДНК в виде спирали. лестница.Остов каждой полинуклеотидной цепи (представлен как лента) состоит из чередующихся остатков фосфата и дезоксирибозы связаны фосфодиэфиром (подробнее …)
Репликация ДНК
Во время репликации бактериального генома каждая нить в двойной спирали ДНК служит шаблоном для синтеза новой комплементарной цепи. Каждая дочь Таким образом, двухцепочечная молекула ДНК содержит одну старую полинуклеотидную цепь и одну вновь синтезированная нить. Такой тип репликации ДНК называется полуконсервативный.Репликация хромосомной ДНК у бактерий начинается с конкретный хромосомный участок, называемый источником, и продолжается двунаправленно, пока процесс завершен (). Когда бактерии делятся бинарным делением после завершения репликации ДНК, реплицированные хромосомы делятся на каждую из дочерних клеток. В области происхождения специфически и временно связываются с клеточной мембраной после того, как репликация ДНК была инициирована, что привело к модели, согласно которой мембрана прикрепление управляет разделением дочерних хромосом (модель репликона).Эти характеристики репликации ДНК во время роста бактерий соответствуют требования генетического материала к точному воспроизведению и наследуется каждой дочерней клеткой во время деления клетки.
Рисунок 5-2
Авторадиография интактной реплицирующейся хромосомы E. coli . Бактерии были радиоактивно помечены тритием тимидином для приблизительно в двух поколениях и подвергались бережному лизированию. Бактериальная ДНК затем был исследован авторадиографией. Вставка показывает репликацию бактериальный (подробнее…)
Экспрессия гена
Генетическая информация, закодированная в ДНК, выражается путем синтеза специфических РНК и белки, и информация течет от ДНК к РНК к белку. ДНК-направленный синтез РНК называется транскрипцией. Поскольку пряди двухспиральные ДНК антипараллельны и комплементарны, только одна из двух цепей ДНК может служат в качестве матрицы для синтеза конкретной молекулы мРНК. Информационные РНК (мРНК) передают информацию от ДНК, и каждая мРНК в бактериях функционирует как матрица для синтеза одного или нескольких конкретных белков.Процесс по нуклеотидная последовательность молекулы мРНК определяет первичный амино кислотная последовательность белка называется трансляцией. Рибосомы, комплексы рибосомные РНК (рРНК) и несколько рибосомных белков, транслируют каждую мРНК в соответствующую полипептидную последовательность с помощью транспортных РНК (тРНК), синтез аминоацил тРНК, факторы инициации и факторы элонгации. Все эти компоненты аппарата для синтеза белка функционируют в производство множества различных белков.Ген — это последовательность ДНК, которая кодирует белок, рРНК или молекула тРНК (продукт гена).
Генетический код определяет, как нуклеотиды в мРНК определяют аминокислоты. в полипептиде. Потому что в мРНК всего 4 разных нуклеотида. (содержащие U, A, C и G), отдельные нуклеотиды не содержат достаточно информации однозначно указать все 20 аминокислот. В динуклеотидах 16 (4 × 4) возможны расположения четырех нуклеотидов, а в Возможны 64 компоновки тринуклеотидов (4 × 4 × 4).Таким образом, требуется минимум три нуклеотида для обеспечения хотя бы одного уникального последовательность, соответствующая каждой из 20 аминокислот. В «Универсальный» генетический код, используемый большинством организмов (), представляет собой тройной код, в котором 61 из 64 возможных тринуклеотида (кодона) кодируют определенные аминокислоты, и любой из три оставшихся кодона (UAG, UAA или UGA) приводят к прекращению перевод. Кодоны, обрывающие цепь, также называются бессмысленными кодонами. потому что они не определяют никаких аминокислот. Генетический код описывается как вырожденный, потому что несколько кодонов могут использоваться для одной аминокислоты, и как неперекрывающиеся, потому что соседние кодоны не имеют общих нуклеотидов.Исключения из «универсального» кода включают использование UGA в качестве кодон триптофана у некоторых видов Mycoplasma и у митохондриальная ДНК и несколько дополнительных кодоновых различий в митохондриальной ДНК из дрожжей , Drosophila и млекопитающих. Трансляция мРНК осуществляется обычно инициируется кодоном AUG для метионина, а соседние кодоны транслируется последовательно, когда мРНК читается с 5 ‘по 3’ направление. Соответствующая полипептидная цепь собирается, начиная с ее аминоконце и продвигаясь к его карбоксиконцу.Последовательность амино кислоты в полипептиде, следовательно, колинеарны с последовательностью нуклеотидов в мРНК и соответствующем гене. Специфические ферментативные реакции участвуют в синтезе ДНК, РНК и белка, выходят за рамки этого глава.
Экспрессия генетических детерминант у бактерий включает однонаправленный поток информации от ДНК к РНК к белку. В бактериофагах либо ДНК, либо РНК. может служить генетическим материалом. При заражении бактерий РНК бактериофаги, молекулы РНК служат матрицами для репликации РНК и мРНК.Исследования ретровирусной группы вирусов животных показывают, что ДНК молекулы могут быть синтезированы из матриц РНК ферментами, обозначенными как РНК-зависимые ДНК-полимеразы (обратные транскриптазы). Этот поворот обычное направление потока генетической информации от РНК к ДНК, а не от ДНК в РНК — важный механизм передачи информации от ретровирусов. закодироваться в ДНК и стать частью геномов животных клетки.
Организация генома
Молекулы ДНК, которые реплицируются как дискретные генетические единицы в бактериях, называются репликоны.У некоторых штаммов Escherichia coli хромосома является в клетке присутствует только репликон. У других бактериальных штаммов есть дополнительные репликоны, такие как плазмиды и бактериофаги.
Хромосомная ДНК
Бактериальные геномы варьируются по размеру от примерно 0,4 × 10 9 до 8,6 × 10 9 дальтон (Да), некоторые из самых маленьких являются обязательными паразиты ( Mycoplasma ) и самые крупные, принадлежащие к бактериям способны к сложной дифференцировке, например, Myxococcus .В количество ДНК в геноме определяет максимальное количество информации, которую он может кодировать. Большинство бактерий имеют гаплоидный геном, единственную хромосому, состоящую из кольцевой двухцепочечной молекулы ДНК. Однако линейные хромосомы имеют был обнаружен у грамположительных Borrelia и Streptomyces spp., Один линейный и один круговой хромосома присутствует в грамотрицательной бактерии Agrobacterium tumefaciens . Единая хромосома общего кишечного бактерия E.coli составляет 3 × 10 9 Да (4500 пары килобаз [kbp]) размером примерно от 2 до 3 процентов от сухой массы клетки. Геном E. coli является всего около 0,1% от размера генома человека, но этого достаточно для кодирования несколько тысяч полипептидов среднего размера (40 кДа или 360 аминокислот).
Хромосома кишечной палочки имеет контурную длину приблизительно 1,35 мм, что в несколько сотен раз длиннее бактериальной клетки, но ДНК суперспиральный и плотно упакованный в бактериальный нуклеоид.Необходимое время для репликации всей хромосомы составляет около 40 минут, что составляет примерно вдвое меньше кратчайшего времени деления этой бактерии. ДНК репликация должна инициироваться так часто, как клетки делятся, чтобы быстро при выращивании бактерий новый виток хромосомной репликации начинается раньше, чем раньше. раунд завершен. При высоких темпах роста может быть четыре хромосомы. реплицируются с образованием восьми во время деления клеток, что связано с завершение раунда хромосомной репликации.Таким образом, хромосома в быстрорастущие бактерии размножаются более чем в одной точке. Репликация хромосомной ДНК бактерий сложна и включает в себя множество различных белки.
Плазмиды
Плазмиды — это репликоны, которые сохраняются как дискретные, внехромосомные генетические элементы в бактериях. Обычно они намного меньше бактериального хромосома, варьирующаяся от менее 5 до более нескольких сотен кб, хотя плазмиды размером до 2 Mbp встречаются у некоторых бактерий. Плазмиды обычно кодируют черты, которые не важны для жизнеспособности бактерий, и воспроизводятся независимо от хромосомы.Большинство плазмид имеют суперспиральную, круговую, двухцепочечные молекулы ДНК, но также были продемонстрированы линейные плазмиды у Borrelia и Streptomyces . Близко связанный или идентичные плазмиды демонстрируют несовместимость; они не могут быть стабильными сохраняется в том же бактериальном хозяине. Классификация плазмид основана на несовместимости или использования конкретных ДНК-зондов в тестах гибридизации с идентифицировать нуклеотидные последовательности, характерные для конкретной плазмиды репликоны.Некоторые гибридные плазмиды содержат более одного репликона. плазмиды кодируют функции, которые способствуют передаче плазмиды от донора бактерии к другим бактериям-реципиентам, но неконъюгативные плазмиды — нет. Конъюгативные плазмиды, которые также способствуют переносу бактериальной хромосомы из бактерии-доноры для других бактерий-реципиентов называются плазмидами фертильности, и обсуждаются ниже. Среднее количество молекул данной плазмиды на бактериальная хромосома называется числом ее копий.Большие плазмиды (> 40 пары килобаз) часто бывают конъюгативными, имеют уменьшенное количество копий (от 1 до нескольких на хромосома), код для всех функций, необходимых для их репликации, и разделяются между дочерними клетками во время деления клеток способом похож на бактериальную хромосому. Плазмиды размером менее 7,5 тысяч пар оснований обычно не сопряжены, имеют большое количество копий (обычно 10–20 на хромосому), полагаются на своего бактериального хозяина, чтобы обеспечить некоторые необходимые функции для репликации и случайным образом распределяются между дочерними клетками на разделение.
Многие плазмиды контролируют важные с медицинской точки зрения свойства патогенных бактерий, включая устойчивость к одному или нескольким антибиотикам, выработку токсинов и синтез структур клеточной поверхности, необходимых для прикрепления или колонизации. Плазмиды, определяющие устойчивость к антибиотикам, часто называют плазмидами R. (или R-факторы). Типичные токсины, кодируемые плазмидами, включают термолабильные и термостойкие энтеротоксины E. coli , эксфолиативный токсин золотистый стафилококк и столбнячный токсин Clostridium tetani .Некоторые плазмиды загадочны и не имеют узнаваемое воздействие на бактериальные клетки, в которых они находятся. Сравнение плазмиды профили — полезный метод оценки возможного родства отдельных клинические изоляты определенного вида бактерий для эпидемиологических исследований. Обсуждается роль плазмид в развитии устойчивости к антибиотикам. ниже.
Бактериофаги
Бактериофаги (бактериальные вирусы, фаги) — инфекционные агенты, которые размножаются как облигатные внутриклеточные паразиты у бактерий.Внеклеточные фаговые частицы метаболически инертны и состоят в основном из белков и нуклеиновой кислоты (ДНК или РНК, но не то и другое вместе). Белки фаговой частицы образуют защитную оболочка (капсид), окружающая плотно упакованный геном нуклеиновой кислоты. Фаг геномы различаются по размеру от примерно 2 до 200 килобаз на цепь нуклеиновой кислоты. кислоты и состоят из двухцепочечной ДНК, одноцепочечной ДНК или РНК. Фаг геномы, как и плазмиды, кодируют функции, необходимые для репликации в бактериях, но в отличие от плазмид они также кодируют белки капсида и неструктурные белки требуется для сборки фага.Несколько морфологически различных типов фага были описаны, в том числе многогранные, нитевидные и сложные. Сложный фаги имеют многогранные головы, к которым хвосты, а иногда и другие придатки (хвост пластины, хвостовые волокна и т. д.).
Один цикл роста фага показан на. Инфекция инициируется адсорбцией фага на специфических рецепторы на поверхности восприимчивых бактерий-хозяев. Капсиды остаются на поверхность клетки, а геномы ДНК или РНК проникают в клетки-мишени (проникновение).Поскольку инфекционность геномной ДНК или РНК намного меньше, чем у зрелых вируса, сразу после заражения наступает период, называемый периодом затмения. во время которого невозможно обнаружить внутриклеточный инфекционный фаг. Заражение РНК или ДНК фага реплицируются, чтобы произвести множество новых копий генома фага, и продуцируются фаг-специфические белки. Для большинства фагов сборки потомства происходит в цитоплазме, а высвобождение потомства происходит путем лизиса клеток. В напротив, нитчатые фаги образуются на клеточной оболочке и высвобождаются не убивая клетки-хозяева.Период затмения заканчивается, когда внутриклеточный появляются заразные потомства. Скрытый период — это интервал от заражения. до появления внеклеточного потомства, а период роста — это интервал от конец латентного периода, пока все фаги не станут внеклеточными. Среднее число частиц фага, продуцируемых каждой инфицированной клеткой, называемого размером взрыва, составляет характерна для каждого вируса и часто составляет от 50 до нескольких сотен. Для обсуждения строения, размножения и классификации животных. вирусы, см. главы 41 и 42.
Рисунок 5-3
Одностадийный рост бактериофага. Культура восприимчивых бактерий синхронно заражается бактериофаг добавлен в момент времени 0 при низкой множественности инфекции. Вскоре после этого неабсорбированный фаг инактивируется добавлением антифаговая антисыворотка, (подробнее …)
Фаги подразделяются на две основные группы: вирулентные и умеренные. Рост вирулентные фаги у чувствительных бактерий разрушают клетки-хозяева. Заражение восприимчивые бактерии к умеренным фагам могут иметь один из двух результатов: литический рост или лизогения.Литический рост умеренных и вирулентных бактериофагов аналогично, что приводит к образованию фагового потомства и гибели бактерий-хозяев. Лизогения — это особый тип латентной вирусной инфекции, при которой геном фага реплицируется как профаг в бактериальной клетке. У большинства лизогенных бактерий гены, необходимые для развития литического фага, не экспрессируются, и производство инфекционный фаг не встречается. Кроме того, лизогенные клетки невосприимчивы к суперинфекция вирусом, который они носят как профаги.Физическое состояние профага не является идентичным для всех вирусов умеренного климата. Например, профаг бактериофага λ в E. coli интегрирован в бактериальную хромосому в определенном месте и реплицируется как часть бактериальная хромосома, тогда как профаг бактериофага P1 у E. coli реплицируется как внехромосомная плазмида.
Рост литического фага происходит спонтанно в небольшой фракции лизогенных клеток, и несколько внеклеточных фагов присутствуют в культурах лизогенных бактерий.Для некоторых лизогенных бактерий синхронная индукция развития литических фагов встречается во всей популяции лизогенных бактерий, когда они обрабатываются агенты, которые повреждают ДНК, такие как ультрафиолет или митомицин С. Потеря профаг из лизогенной бактерии, переводя его в нелизогенное состояние и восстановление восприимчивости к инфекции первоначально присутствующим фагом как профаг, называется лечением.
Некоторые фаги умеренного пояса содержат гены бактериальных характеристик, которые не связаны с развитием литического фага или лизогенным состоянием, и экспрессия такие гены называют фаговой конверсией (или лизогенной конверсией).Примеры конверсия фагов, которая важна для микробной вирулентности, включает производство токсина дифтерии Corynebacterium diphtheriae , эритрогенный токсин Streptococcus pyogenes (группа А β-гемолитические стрептококки), ботулинический токсин от Clostridium botulinum , а шига-подобные токсины — E. coli . В В каждом из этих примеров ген, кодирующий бактериальный токсин, присутствует в умеренный геном фага. Специфичность O-антигенов в Salmonella также можно контролировать с помощью фаговой конверсии.Типирование фагов — это тестирование штаммов определенного вида бактерий на восприимчивость к специфическим бактериофагам. Паттерны восприимчивости к набор типирующих фагов предоставляет информацию о возможном родстве отдельные клинические изоляты. Такая информация особенно полезна для эпидемиологические исследования.
Мутация и отбор
Мутации — это наследуемые изменения генома. Спонтанные мутации у человека бактерии встречаются редко. Некоторые мутации вызывают изменения фенотипических характеристик; то о возникновении таких мутаций можно судить по вызываемым ими эффектам.В контрольные организмы, специфичные для микробной генетики, обозначены как штаммы дикого типа, а потомки, в геномах которых есть мутации, называются мутантами. Таким образом, мутанты характеризуются унаследованными различиями между ними и их предковые штаммы дикого типа. Вариантными формами конкретной генетической детерминанты являются: называются аллелями. Генотипические символы — строчные буквы, сокращения выделены курсивом. указать отдельные гены с надстрочным индексом (+), указывающим на дикий тип аллель. Фенотипические символы пишутся с большой буквы, а не курсивом, чтобы их можно было различить. от генотипических символов.Например, генотипический символ способности производить β-галактозидаза, необходимая для ферментации лактозы, является lacZ + и мутанты, которые не могут производить β-галактозидазы lacZ . Фенотип ферментации лактозы обозначается Lac + , а неспособность ферментировать лактозу Lac — .
Обнаружение мутантных фенотипов
Селективные и дифференциальные среды полезны для выделения бактериальных мутантов. Некоторые селективные среды позволяют расти определенным мутантам, но не позволяют штаммы дикого типа для выращивания.Редкие мутанты можно выделить с помощью таких селективных СМИ. Дифференциальные среды позволяют бактериям дикого типа и мутантным бактериям расти и формироваться. колонии, различающиеся по внешнему виду. Обнаружение редких мутантов по дифференциалу media ограничено общим количеством колоний, которые можно наблюдать. Рассматривать штамм E. coli дикого типа, чувствительный к антибиотик стрептомицин (фенотип Str s ) и может использовать лактозу в качестве единственный источник углерода (фенотип Lac + ).Спонтанно Встречающиеся мутанты Str r редки и обычно встречаются с частотами менее одной на 109 бактерий в культурах дикого типа E. coli . Тем не менее, мутанты Strr можно легко выделить, используя селективная среда, содержащая стрептомицин, поскольку Str s дикого типа бактерии погибают. Выделение лактозонегативных (фенотип Lac — ) мутанты E. coli представляют собой другая проблема. На минимальных средах с лактозой в качестве единственного источника углерода, Lac + штаммы дикого типа будут расти, но Мутанты Lac — не могут расти.На дифференциальных средах, таких как МакКонки-лактозный агар или эозин-метиленовый синий-лактозный агар, Lac + дикого типа и мутантные штаммы Lac — E. coli можно отличить по цвету, но самопроизвольно Мутанты Lac — слишком редки, чтобы их легко выделить. Селективная среда для мутантов Lac — E. coli можно получить путем включения химических аналогов лактозы, которые превращаются в токсичные метаболиты бактериями Lac + , но не мутантами Lac —.Мутанты Lac — затем могут расти на такой среде, но бактерии дикого типа Lac + убиты.
Мутации, инактивирующие основные гены гаплоидных организмов, обычно летальные, но такие потенциально смертельные мутации часто можно изучить, если их экспрессия контролируется манипулированием экспериментальными условиями. Для Например, мутация, которая увеличивает термолабильность важного гена продукт может предотвратить рост бактерий при 42 ° C, хотя мутант бактерия все еще может расти при 25 ° C.И наоборот, чувствительные к холоду мутанты выражают мутантный фенотип при низкой температуре, но не при высокой температуре. Чувствительные к температуре и холоду мутации являются примерами условных мутации, как и подавляемые мутации, описанные далее в этой главе. А условный летальный фенотип указывает на то, что мутантный ген важен для жизнеспособность.
Спонтанные и индуцированные мутации
Скорость мутации у бактерий определяется точностью репликации ДНК, возникновение повреждения ДНК и эффективность механизмов восстановления поврежденной ДНК.
Для конкретного бактериального штамма в определенных условиях роста мутация скорость для любого конкретного гена постоянна и выражается как вероятность мутации на деление клетки. В популяции бактерий, выросших из небольшого inoculum, доля мутантов обычно прогрессивно увеличивается по мере увеличения размера бактериальной популяции увеличивается.
Мутации у бактерий могут происходить спонтанно и независимо от для их обнаружения используются экспериментальные методы. Этот принцип был впервые продемонстрирован тестом на колебания ().В количество устойчивых к фагам мутантов E. coli в повторностях культуры, выращенные из небольших инокул, были измерены и сравнены с культурами, выращенными в несколько образцов, взятых из одной культуры. Если мутации по устойчивости к фагам произошла только после воздействия фага, вариабельность количества мутантов между культурами должны быть одинаковыми в обоих наборах условий. Напротив, если устойчивые к фагам мутанты возникли спонтанно до воздействия от бактерий к фагу, количество мутантов должно быть более вариабельным в самостоятельно выращенные культуры, поскольку различия в размерах бактерий популяции, когда появился первый мутант, будет способствовать наблюдаемому изменчивость.Данные показали, что мутации устойчивости к фагам в E. coli возник спонтанно с постоянной вероятностью на деление клетки.
Рисунок 5-4
Испытание на колебания. Различия в количестве колоний устойчивых к фагам мутантов в повторные образцы из одной субкультуры были небольшими и отражали только ожидаемые колебания из-за ошибок выборки. Напротив, количество устойчивых к фагам колоний (подробнее …)
Посев на реплики подтвердил, что мутации у бактерий могут возникать спонтанно, без воздействия на бактерии селективных агентов ().Для покрытия реплик: плоский стерильный вельветовый поверхность используется для сбора инокулята с поверхности эталонной чашки с агаром и перенесите образцы в другие чашки с агаром. Таким образом, образцы Популяция бактерий с мастер-планшета переносится на реплики не искажая их пространственного расположения. Если реплики содержат селективной среды и мастер-планшетов нет, положения отобранных мутантов можно отметить колонии на чашках-репликах и бактерии, которые не подвергались воздействию к условиям отбора могут быть изолированы с тех же позиций на мастер пластина.Мутанты E. coli , устойчивые к бактериофагу Т1 или стрептомицину были выделены таким образом, не подвергая бактерии дикого типа к бактериофагу или антибиотику.
Рисунок 5-5
Выявление ранее существовавших бактериальных мутантов по реплике покрытие. Мастер-планшет был сильно засеян образцом из чистых культур чувствительные к фагам бактерии. После инкубации бактерии от хозяина планшеты были перенесены путем посева реплик в дубликаты (подробнее …)
На частоту мутаций влияют как экологические, так и генетические факторы.Контакт с бактерий в мутагенные агенты вызывает увеличение скорости мутаций, иногда на на несколько порядков. Многие химические и физические агенты, включая рентгеновские лучи и ультрафиолет, обладают мутагенной активностью. Канцерогенные химические вещества для животных часто являются мутагенными для бактерий или могут быть преобразованы животными ткани на метаболиты, которые являются мутагенными для бактерий. Стандартизированные тесты для мутагенность бактерий используются в качестве скрининговых процедур для выявления агенты окружающей среды, которые могут быть канцерогенными для человека.Гены-мутаторы в бактерии вызывают увеличение частоты спонтанных мутаций для самых разных другие гены. Экспрессия этих генов, вызванная повреждением ДНК (см. SOS-ответ позже), позволяет восстанавливать повреждения ДНК, которые в противном случае были бы летальными, но за счет подверженный ошибкам механизм, который увеличивает скорость мутации. В целом скорость мутации — вероятность того, что мутация произойдет где-то в бактериальный геном на деление клетки — относительно постоянен для множество организмов с геномами разного размера и, по-видимому, важный фактор в определении пригодности бактериального штамма к выживанию в природе.Большинство мутаций вредны, и существует риск неблагоприятных мутаций для отдельные бактерии должны быть сбалансированы с положительным значением изменчивости, поскольку механизм адаптации бактериальных популяций к изменяющимся условиям окружающей среды. условия.
Молекулярная основа мутаций
Мутации классифицируются на основе структурных изменений, происходящих в ДНК (). Некоторые мутации локализованы в коротких сегментах ДНК (например, при нуклеотидных заменах, микроделеции и микровставки).Другие мутации затрагивают большие области ДНК и включают делеции, вставки или перестройки сегментов ДНК.
Когда нуклеотидная замена происходит в области ДНК, которая кодирует полипептид, один из трех нуклеотидов в одном кодоне соответствующая молекула мРНК будет изменена. Тихие мутации не меняют структура или функция полипептида, потому что один кодон в мРНК заменен на другой для той же аминокислоты. Другие замены вызывают замену одной аминокислоты. заменен другим в конкретном положении в полипептиде соответствующий измененному кодону.Мутации, приводящие к замене одного аминокислота для другой в полипептидной цепи называется миссенс-мутациями. Влияние аминокислотных замен на функцию полипептидного гена продукт различается и зависит от местоположения и идентичности аминокислоты замена. Мутантные полипептиды, содержащие замены аминокислот, обычно имеют общие антигенные детерминанты с полипептидом дикого типа и часто имеют некоторые остаточная биологическая активность. Мутации, приводящие к замене аминокислоты кислотный кодон с терминирующим кодоном называют нонсенс-мутациями.Это результаты в продукции амино-концевого фрагмента нормального полипептида, когда мутантная мРНК транслируется. Бессмысленные мутации часто приводят к полной потере активность генного продукта.
Из-за триплетной природы генетического кода последствия мутаций вызванные вставками или удалениями небольшого количества нуклеотидов (микровставки, микроделеции) зависят как от количества, так и от последовательности вовлеченные нуклеотиды. Удаление или добавление кратных трех нуклеотидов пары не влияют на рамку считывания, но вызывают удаление или добавление соответствующее количество аминокислот в одном сайте полипептида.Если новый кодон обрыва цепи вводится, преждевременное обрыв цепи происходит в пределах полипептид. Напротив, добавление или удаление других номеров пары нуклеотидов изменяют рамку считывания для всего сегмента мРНК от мутация дистального конца гена. Следовательно, мутации сдвига рамки считывания может вызвать резкие изменения в структуре и активности полипептида генные продукты, и их часто классифицируют как бессмысленные мутации.
Тесты комплементации
Чтобы определить, находятся ли мутации в одном и том же гене или в разных генах, тесты комплементации проводятся с частично диплоидными бактериальными штаммами ().Две копии региона бактериальной хромосомы, несущей мутацию, присутствуют в том же бактерия, причем каждая копия содержит разные мутации (мутации находятся в транс расположение). Фенотип дикого типа указывает на то, что мутации разные гены. Это явление называется комплементацией. Если мутант фенотипа, необходимо провести контрольный эксперимент с мутации в цис-расположении, чтобы исключить возможность того, что дикий тип аллели не могут нормально экспрессироваться в частично диплоидном штамме бактерий.Изначально тесты комплементации назывались «цис-транс». тесты, а термин цистрон иногда используется как синоним гена. Дополнительные тесты могут быть выполнены и интерпретированы, даже если биохимические функции продуктов гена неизвестны.
Рисунок 5-6
Комплементация — это метод проверки функционального гена товары. Два мутанта со схожими фенотипами (неспособность конвертировать субстрат X к продукту Z). Мутации в этих штаммах обозначены a и b, соответственно, и аллели дикого типа (подробнее…)
В качестве примера рассмотрим использование теста дополнения для характеристики двух независимо полученные мутанты Lac — E. coli . Биохимический путь утилизации лактозы требует β-галактозид пермеаза (генотипический символ lac Y) для транспортировки лактоза в бактериальную клетку и β-галактозидазу (генотипический символ lacZ ) для превращения лактозы в D-глюкозу и D-галактозу. Мутанты без β-галактозид пермеазы или β-галактозидазы не могут используйте лактозу для роста.Если мутации в обоих Lac — мутанты инактивировали тот же белок (например, β-галактозид), затем частичный диплоидный штамм, содержащий генов lacZ от обоих мутанты в транс-структуре не могут использовать лактозу. В напротив, если бы генотипы двух мутантов были lacZ + lacY и lacZ lacY + , частично диплоидная бактерия будет производить активную β-галактозидазу из lacZ + определитель и активный β-галактозид пермеаза из лаковый + определитель.Дополнение произойдет, и частично диплоидный штамм будет использовать лактозу.
Реверсия и подавление
Мутации, которые преобразуют фенотип из дикого типа в мутантный, называются прямыми мутации и мутации, которые изменяют фенотип с мутантного обратно на дикий тип называются обратными мутациями (реверсиями). Бактериальные штаммы, содержащие обратные мутации называются ревертантами. Анализ мутаций, вызывающих фенотипическая реверсия дает полезную информацию. Обратные мутации, восстанавливающие точные нуклеотидные последовательности ДНК дикого типа являются истинными реверсиями.Истинный ревертанты идентичны штаммам дикого типа как генотипически, так и фенотипически. Обратные мутации, которые не восстанавливают точный нуклеотид Последовательности ДНК дикого типа называются супрессорными мутациями (супрессорами). Некоторые ревертанты, несущие супрессорные мутации, фенотипически неотличимы от штаммов дикого типа. Другие ревертанты, называемые псевдоревертанты, фенотипически отличимые от штаммов дикого типа, например, из-за тонких различий в характеристиках ферментативного восстановленная активность (например, удельная активность, субстрат специфичность, кинетические константы или восприимчивость к термическому или химическому инактивация).Распознавание псевдоревертантных фенотипов предполагает наличие супрессорных мутаций.
Супрессорные мутации могут быть внутригенными или экстрагенными. Внутригенные супрессоры расположены в том же гене, что и подавляемые ими прямые мутации. В возможные места и характер внутригенных супрессоров определяются исходная прямая мутация и отношения между первичной структурой продукта гена и его биологической активности. Экстрагенные супрессоры находятся в разных генах от мутаций, эффекты которых они подавляют.В способность экстрагенных супрессоров подавлять множество независимых мутаций можно протестировать. Некоторые экстрагенные супрессоры специфичны для определенных генов, некоторые специфичны для определенных кодонов, а некоторые имеют другую специфичность узоры. Экстрагенные супрессоры, которые меняют фенотипические эффекты кодоны, завершающие цепь, были хорошо охарактеризованы, и было обнаружено, что они изменяют структура специфических тРНК. Конкретная супрессорная тРНК может разрешать определенные кодон обрыва цепи, который будет транслироваться, что приведет к включению специфической аминокислоты в формирующийся полипептид в положении, соответствующем к кодону обрыва цепи.У бактерии, имеющей оканчивающуюся цепь мутация и соответствующий экстрагенный супрессор, трансляция мРНК содержащий мутантный кодон, следовательно, может привести к образованию полноразмерного полипептид. Биологическая активность полноразмерного полипептида, образованного как Последствия подавления зависят как от количества вырабатываемого белка, так и от функциональные последствия замены конкретной аминокислоты, определяемые супрессорная тРНК.
Обмен генетической информацией
Биологическое значение сексуальности у микроорганизмов заключается в увеличении вероятность того, что редкие независимые мутации произойдут вместе в одном микробе и подвергнуться естественному отбору.Генетические взаимодействия между микробами позволяют их геномы эволюционируют гораздо быстрее, чем за счет одной лишь мутации. Представитель явления, имеющие медицинское значение, которые включают обмен генетической информацией или геномные перестройки включают быстрое появление и распространение антибиотика. плазмиды устойчивости, вариации жгутиковых фаз у сальмонелл и антигенные изменение поверхностных антигенов у Neisseria и Borrelia.
Половые процессы у бактерий включают передачу генетической информации от донора к реципиент и приводит либо к замене аллелей донора на аллели реципиента или добавление донорских генетических элементов в геном реципиента.Трансформация, трансдукция и конъюгация — это половые процессы, в которых используются разные механизмы для ввести донорскую ДНК в реципиентные бактерии (). Поскольку донорская ДНК не может сохраняться в бактерии-реципиенте, если это часть репликона, рекомбинация между геномами донора и реципиента часто требуется для получения стабильного гибридного потомства. Наиболее вероятна рекомбинация когда донорская и реципиентная бактерии происходят от одного или близкородственного происхождения разновидность.
Рисунок 5-7
Обмен генетической информацией в бактериях.Трансформация, трансдукция и конъюгация различаются способами введение ДНК из клетки-донора в клетку-реципиент. А) В трансформация, отбираются фрагменты ДНК, освобожденные от донорских бактерий вверх по компетентному (подробнее …)
Для того, чтобы рекомбинант был обнаружен, его фенотип должен отличаться от обоих родительских фенотипы. Перед рекомбинантным фенотипом может потребоваться рост или деление клеток. выражается. Задержка экспрессии рекомбинантного фенотипа до гаплоида сегрегация рекомбинантного генома называется задержкой сегрегации и задержкой до тех пор, пока синтез продуктов, кодируемых донорскими генами, называется фенотипическим отставанием.Тестирование на сцепление (неслучайная перегруппировка родительских аллелей в рекомбинантных потомство) возможно, когда родительские бактерии имеют разные аллели для нескольких гены. Донорский аллель невыбранного гена с большей вероятностью присутствует в рекомбинантов, если он связан с выбранным донорским геном, чем если он не связан с выбранный донорский ген. Количественный анализ связей позволяет построить генетические карты. Геном E. coli является кольцевым (), что определяется как генетическими сцепление и прямой биохимический анализ хромосомной ДНК, и генетическая карта колинеарно с физической картой хромосомной ДНК.Генетическое и физическое картирование также используются для анализа внехромосомных репликонов, таких как бактериофаги и плазмиды.
Рисунок 5-8
Круговая генетическая карта E. coli . Позиции репрезентативных генов указаны на внутреннем круге. Расстояния между генами калибруются в минутах, в зависимости от времени. требуется для переноса при конъюгации. Положение треонина (th) локус произвольно (подробнее …)
Многие бактерии имеют системы рестрикции-модификации (РМ), состоящие из модифицирующих ферменты, которые метилируют остатки аденина или цитозина в определенных последовательностях в их собственная ДНК и соответствующие эндонуклеазы рестрикции, которые расщепляют чужеродную ДНК, которая не несет специфической модификации в тех же целевых последовательностях.Некоторые рестрикционные ферменты будут расщеплять только ДНК, метилированную в определенных последовательности. Эти системы ограничения, которые могли развиться для защиты бактерий против вторжения фагов или плазмид, являются важным барьером для генетических обмены между различными бактериальными штаммами или видами. Недавние данные свидетельствуют о что плазмидные системы RM могут быть способом для плазмиды обеспечить ее перенос в штамм-хозяин, поскольку клетки, теряющие плазмиду (и соответствующие защитные ген метилазы) убиваются под действием более стабильного рестрикционного фермента, который атакует недавно реплицированную, но неизмененную хромосомную ДНК.
Трансформация
При трансформации захватываются фрагменты ДНК, высвобожденные из донорских бактерий. непосредственно из внеклеточной среды бактериями-реципиентами. Рекомбинация происходит между отдельными молекулами трансформирующей ДНК и хромосомами реципиентные бактерии. Чтобы быть активными в трансформации, молекулы ДНК должны находиться на длиной не менее 500 нуклеотидов, и трансформирующая активность быстро разрушается обработкой ДНК дезоксирибонуклеазой. Молекулы трансформирующей ДНК соответствуют к очень маленьким фрагментам бактериальной хромосомы.Котрансформация генов поэтому маловероятно, если они не связаны настолько тесно, что могут быть кодируется на одном фрагменте ДНК. Трансформация была обнаружена в Streptococcus pneumoniae и встречается у других бактериальных родов, в том числе Haemophilus , Neisseria , Bacillus и Staphylococcus . Способность бактерий для поглощения внеклеточной ДНК и трансформации, называемой компетентность зависит от физиологического состояния бактерий.Многие бактерии которые обычно некомпетентны, могут быть использованы для анализа ДНК в лаборатории манипуляции, такие как кальциевый шок или воздействие высоковольтного электрического пульс (электропорация). У некоторых бактерий (включая Haemophilus и Neisseria) Поглощение ДНК зависит от наличия определенных олигонуклеотидных последовательностей в трансформирует ДНК, но у других (включая Streptococcus pneumoniae) захват ДНК не зависит от последовательности. Компетентные бактерии также могут занимать неповрежденные ДНК бактериофага (трансфекция) или плазмидная ДНК, которая затем может реплицироваться как внехромосомные генетические элементы в бактериях-реципиентах.Напротив, фрагмент хромосомной ДНК от бактерии-донора обычно не может реплицироваться в бактерия-реципиент, если она не станет частью репликона путем рекомбинации. Исторически сложилось так, что характеристика «трансформирующего принципа» из S. pneumoniae при условии, что первое прямое доказательство наличия ДНК генетический материал.
Трансдукция
При трансдукции бактериофаги действуют как векторы для введения ДНК от донора бактерии в бактерии-реципиенты путем инфицирования. Для некоторых фагов, называемых генерализованные трансдуцирующие фаги, небольшая фракция вирионов, продуцируемых во время литический рост аберрантный и содержит случайный фрагмент бактериального генома вместо фаговой ДНК.Каждый индивидуальный трансдуцирующий фаг несет свой набор тесно связанных генов, представляющих небольшой сегмент бактериального генома. Трансдукция, опосредованная популяциями таких фагов, называется генерализованной. трансдукция, потому что каждая часть бактериального генома имеет приблизительно такая же вероятность передачи от донора к бактериям-реципиенту. Когда генерализованный трансдуцирующий фаг заражает реципиентную клетку, экспрессия передаются донорские гены. Абортивная трансдукция относится к временным экспрессия одного или нескольких донорских генов без образования рекомбинантного потомства, тогда как полная трансдукция характеризуется образованием стабильных рекомбинанты, которые наследуют донорские гены и сохраняют способность их экспрессировать.В прерванная трансдукция донорский фрагмент ДНК не реплицируется, и среди потомство исходного трансдуктанта только одна бактерия содержит донорскую ДНК фрагмент. У всех остальных потомков продукты донорских генов постепенно становятся разбавляется после каждого поколения роста бактерий до тех пор, пока донорский фенотип не сможет больше не выражаться. На селективной среде, на которой только бактерии с донорский фенотип может расти, абортивные трансдуктанты производят крошечные колонии, которые легко отличить от колоний стабильных трансдуктантов.Частота абортивной трансдукции обычно на один-два порядка больше чем частота генерализованной трансдукции, что указывает на то, что большинство клеток инфицированные генерализованными трансдуцирующими фагами, не продуцируют рекомбинантные потомство.
Специализированная трансдукция отличается от обобщенной трансдукции несколькими способами. Это опосредуется только определенными умеренными фагами и лишь несколькими конкретными донорами. гены могут передаваться бактериям-реципиентам. Специализированные трансдуцирующие фаги образуются только тогда, когда лизогенные донорские бактерии входят в литический цикл и высвобождают фаговое потомство.Специализированные фаги-трансдукторы — это редкие рекомбинанты, которые не имеют части нормального генома фага и содержат часть бактериального хромосома расположена рядом с местом прикрепления профага. Многие специализированные трансдуцирующие фаги являются дефектными и не могут завершить литический цикл фага рост в инфицированных клетках, если не присутствуют фаги-помощники, функции фага. Специализированная трансдукция является результатом лизогенизации бактерия-реципиент специализированным трансдуцирующим фагом и экспрессией донорские гены.Конверсия фагов и специализированная трансдукция имеют много сходства, но происхождение генов конверсии в умеренном климате фаги неизвестны.
Конъюгация
При конъюгации прямой контакт между донорскими и реципиентными бактериями приводит к установление цитоплазматического мостика между ними и перенос части или всего донорского генома реципиенту. Донорская способность определяется конкретными конъюгативные плазмиды называются плазмидами фертильности или половыми плазмидами.
Плазмида F (также называемая фактором F) E.coli — прототип для плазмид фертильности грамотрицательных бактерий. Штаммы E. coli с внехромосомной плазмидой F называются F + и функционируют как доноры, тогда как штаммы, лишенные Плазмиды F представляют собой F — и ведут себя как реципиенты. Спряжение функции плазмиды F определяются кластером из не менее 25 переносов ( tra ) гены, определяющие экспрессию F-пилей, синтез и перенос ДНК во время спаривания, вмешательство в способность F + бактерии для выполнения и других функций.Каждая бактерия F + имеет от 1 до 3 пилей F, которые связываются с определенным белок внешней мембраны (продукт гена ompA) на бактериях-реципиентах, чтобы инициировать вязка. Образуется межклеточный цитоплазматический мостик, и одна нить F плазмидная ДНК передается от донора к реципиенту, начиная с уникального источника и прогрессирует в направлении от 5 ‘до 3’. Переданный цепь превращается в кольцевую двухцепочечную плазмидную ДНК F у реципиента бактерии, и у донора синтезируется новая цепь для замены перенесенная прядь.Обе экзонъюгантные бактерии — F + , поэтому плазмида F может распространяться инфекцией среди генетически совместимых популяции бактерий. Помимо роли F-пилей в конъюгации, они также функционируют как рецепторы для донор-специфичных (специфичных для мужчин) фагов.
Плазмида F в E. coli может существовать как внехромосомная генетический элемент или быть интегрированным в бактериальную хромосому (). Поскольку плазмида F и Бактериальные хромосомы представляют собой обе кольцевые молекулы ДНК, взаимная рекомбинация между ними образуется более крупный круг ДНК, состоящий из вставленной плазмидной ДНК F линейно в хромосому. E. coli содержит несколько копий нескольких различных генетических элементов, называемых последовательностями вставки (см. транспозоны для более подробной информации), в различных местах в его хромосоме и в Плазмида F. Гомологичная рекомбинация между последовательностями вставки в хромосома и плазмида F приводит к преимущественной интеграции плазмиды F на хромосомных участках, где расположены последовательности вставки. Хромосомный сайты, где обнаруживаются последовательности вставки, различаются, однако, среди штаммов E.coli .
Рисунок 5-9
Роль плазмиды F в определении состояний донора и реципиента E. coli . Плазмида F представляет собой специфические конъюгативные плазмиды, которые контролировать донорскую способность E. coli . F- штаммы отсутствуют плазмиды F и являются генетическими реципиентами. F + штаммы гавани (подробнее …)
Штамм E. coli с интегрированной плазмидой F сохраняет свою способность выступать донором в супружеских отношениях. Поскольку донорский штамм с интегрированные факторы F могут передавать хромосомные гены реципиентам с высоким КПД, они называются Hfr ( H igh f Requency r ecombination) штаммов.Перенос одноцепочечной ДНК от донора Hfr к реципиенту начинается от источника внутри плазмиды F и происходит, как описано выше, за исключением того, что перенесенная ДНК является гибридной. репликон, состоящий из плазмиды F, интегрированной в бактериальную хромосому. Перенос всего этого репликона, включая бактериальную хромосому, требует примерно 100 минут. Идентичность первого хромосомного гена, который будет переносится и полярность хромосомного переноса определяется сайтом интеграции плазмиды F и ее ориентации относительно бактериальная хромосома.Потому что спаривающиеся бактерии обычно отделяются спонтанно до того, как вся хромосома будет перенесена, конъюгация обычно переносится только фрагмент донорской хромосомы в реципиент. Вероятность того, что донорский ген войдет в бактерию-реципиент во время уменьшения конъюгации, следовательно, как его расстояние от начала F (и, следовательно, время его передача) увеличивается. Спаривающиеся клетки также могут быть разделены экспериментально с помощью подвергая их сильным сдвигающим усилиям в механическом блендере; это называется прерванной вязкой.Формирование рекомбинантного потомства требует рекомбинация между перенесенной донорской ДНК и геномом реципиента бактерия. Анализ потомства от вязок, прерванных после разных интервалы демонстрируют, какие хромосомные гены передаются первыми конкретных донорских штаммов, последовательное время входа для генов, которые передаются впоследствии, и все более низкая вероятность того, что гены перенесенные позже появятся в рекомбинантном потомстве. Круглость генетическая карта E.coli был первоначально выведен из перекрывающиеся, циклически переставленные группы связанных генов, которые были перенесены рано отдельными донорскими штаммами, в которых фактор F был интегрирован на разные хромосомные места.
При скрещивании бактерий F + и F — , только плазмида F передается реципиентам с высокой эффективностью. Хромосомные гены передаются с очень низкой эффективностью, и это спонтанные мутанты Hfr в популяциях F + , которые опосредуют передача донорских хромосомных генов.В вязках между Hfr и F — штаммы , сегмент плазмиды F, содержащий tra область переносится последней, после всего бактериального хромосома перенесена. Большинство рекомбинантов от вязок между Hfr и F — клетки не могут наследовать весь набор плазмиды F. гены и фенотипически являются F —. В вязках между F + и F — штаммы , плазмида F быстро распространяется среди бактериальной популяции, и большинство рекомбинантов являются Факс + .
Интегрированные плазмиды F в штаммах Hfr иногда могут быть вырезаны из бактериального хромосома. Если вырезание полностью обращает процесс интегрирования, F + клетки производятся. Однако в редких случаях иссечение происходит путем рекомбинации с участием инсерционных последовательностей или других генов на бактериальном хромосомы, расположенные на некотором расстоянии от исходного сайта интеграции. В таких случаях сегменты бактериальной хромосомы могут включаться в гибридные плазмиды F, которые называют плазмидами F ‘(см.).Подобными процессами сегменты бактериального хромосома иногда может быть включена в плазмиды R с образованием гибридных R ‘плазмиды. Конъюгативные плазмиды R ‘могут функционировать как плазмиды фертильности, потому что они могут интегрироваться в бактериальную хромосому путем гомологичной рекомбинации и опосредуют передачу хромосомных генов во время спаривания с бактериями-реципиентами. F ‘ плазмиды, R ‘плазмиды, специализированные трансдуцирующие фаги и рекомбинантные плазмиды или фаги, сконструированные путем клонирования генов (описанные ниже), представляют собой гибридные репликоны которые могут включать сегменты бактериальной хромосомы.Следовательно, любой из этих генетические элементы могут быть использованы для создания частично диплоидных бактериальных штаммы, необходимые для тестов на комплементарность и других целей.
Конъюгация также происходит у грамположительных бактерий. Грамположительные донорские бактерии вырабатывают адгезины, которые заставляют их агрегироваться с реципиентными клетками, но половые пили не участвуют. У некоторых видов Streptococcus , реципиент бактерии производят внеклеточные половые феромоны, которые вызывают изменение фенотипа донора. экспрессироваться бактериями, несущими соответствующую конъюгативную плазмиду, и конъюгативная плазмида не дает донорским клеткам продуцировать соответствующие феромон.
Рекомбинация
Рекомбинация включает разрыв и соединение родительских молекул ДНК с образованием гибридные, рекомбинантные молекулы. Было обнаружено несколько различных видов рекомбинации. идентифицированы, которые зависят от различных характеристик участвующих геномов и требуют активности различных генных продуктов. Специфические ферменты, которые действуют на ДНК (например, экзонуклеазы, эндонуклеазы, полимеразы, лигазы) участвуют в рекомбинации. Подробное обсуждение биохимических событий в рекомбинации выходит за рамки данной главы.
Обобщенная рекомбинация включает молекулы донорной и реципиентной ДНК, которые имеют гомологичные нуклеотидные последовательности. Взаимные обмены могут происходить между любыми гомологичные сайты-доноры и реципиенты. В E. coli продукт гена recA важен для генерализованной рекомбинации, но и другие генные продукты тоже участвуют.
Сайт-специфическая рекомбинация включает взаимные обмены только между определенными сайты в молекулах донорской и реципиентной ДНК. Ген recA продукт не требуется для сайт-специфической рекомбинации.Интеграция умеренный бактериофаг l в хромосому E. coli хорошо изученный пример сайт-специфической рекомбинации (). Конкретные сайты вложений (att) на Хромосома E. coli и ДНК фага λ имеют общие основная последовательность из 15 нуклеотидов, внутри которой происходит реципрокная рекомбинация, фланкированы соседними последовательностями, не гомологичными фаговым и бактериальным геномы. В фаге λ продукт гена int (интеграза) требуется для специфичного для сайта интеграционного события в лизогенизация; продукты int и Гены xis (эксцизионаза) необходимы для дополнительное сайт-специфическое иссечение, которое происходит во время индукции литического развитие фага в лизогенных клетках.
Рисунок 5-10
Интеграция и удаление бактериофага λ примеры сайт-специфической рекомбинации. λ ДНК показана тонкими линиями, а хромосомная ДНК — толстыми линиями. линий. Места прикрепления (атт) представляют собой закрытые ящики для бактериального хромосома и открытые боксы (подробнее …)
Незаконная рекомбинация — это термин, используемый для описания негомологичных, аберрантных рекомбинационные события, такие как те, которые участвуют в формировании специализированных трансдуцирующие фаги. Механизмы нелегитимной рекомбинации: неизвестный.
Транспозоны
Транспозоны — это сегменты ДНК, которые могут перемещаться из одного участка молекулы ДНК в другие сайты-мишени в той же или другой молекуле ДНК. Процесс называется транспозицией и происходит с помощью механизма, который не зависит от обобщенная рекомбинация. Транспозоны являются важными генетическими элементами, потому что они вызывают мутации, опосредуют геномные перестройки, функционируют как переносимые области генетической гомологии, приобретают новые гены и вносят свой вклад в их распространение среди бактериальных популяций.Часто введение транспозона прерывает линейную последовательность гена и инактивирует его. Транспозоны имеют важную роль в возникновении делеций, дупликаций и инверсий сегментов ДНК, как а также слияния репликонов. Транспозоны не являются самовоспроизводящимися генетическими элементы, однако, и они должны интегрироваться в другие репликоны для поддержания стабильно в бактериальных геномах.
Большинство транспозонов имеют ряд общих черт. Каждый транспозон кодирует функции, необходимые для его транспозиции, в том числе фермент транспозазы, который взаимодействует со специфическими последовательностями на концах транспозона.В течение при транспозиции короткая последовательность целевой ДНК дублируется, и транспозон вставлен между непосредственно повторяющимися целевыми последовательностями. Длина этого короткая дупликация варьирует, но характерна для каждого транспозона. В Предполагается, что дупликация включает асимметричное расщепление ДНК в мишени. сайт с последующим синтезом новых комплементарных цепей, соответствующих область между сайтами расщепления. Некоторые транспозоны вставляются практически в любые целевая последовательность, тогда как другие имеют относительно строгую целевую специфичность.Различают два типа транспозиции. Удаление транспозона из донорский сайт с последующей его вставкой в целевой сайт называется нерепликативным. транспонирование. Если транспозон на донорском сайте реплицируется и копия вставлен в целевой сайт, однако процесс называется репликативным транспозиция. Процесс репликативной транспозиции может включать образование коинтеграция, одиночная кольцевая молекула ДНК, состоящая из двух репликонов, соединенных с копиями транспозона в чередующейся последовательности.Резолюция коинтеграция в составляющие его репликоны часто достигается за счет кодируемая транспозоном резольваза, которая катализирует сайт-специфическую рекомбинацию между транспозоны. Обобщенная рекомбинация между гомологичными транспозонами может также приводят к образованию или расслоению коинтегратов. Транспозиция отличается от сайт-специфической рекомбинации путем дублирования сегмента целевой последовательности и с использованием множества различных целевых последовательностей для одного донора последовательность.
Большинство транспозонов в бактериях можно разделить на три основных класса ().Последовательности вставки и связанные составные транспозоны составляют первый класс. Последовательности вставки самые простые в структуре и кодируют только функции, необходимые для транспонирования. Известный Последовательности вставки варьируются по длине примерно от 780 до 1500 нуклеотидов. пары, имеют короткие (15-25 пар оснований) перевернутые повторы на концах и не являются тесно связаны друг с другом. ДНК между инвертированными концевыми повторами содержит один (или редко два) гена транспозазы и не кодирует резольвазу.Сложные транспозоны различаются по длине от примерно 2000 до более чем 40000. пары нуклеотидов и содержат последовательности вставки (или близкородственные последовательности) на каждом конце, обычно в виде перевернутых повторов. Весь сложный элемент может транспонировать как единое целое. ДНК между концевыми вставками последовательностей комплекса транспозоны кодируют несколько функций, которые не важны для транспозиции. У важных с медицинской точки зрения бактерий гены, определяющие приверженность к лечению. антигенов, токсинов или других факторов вирулентности, или указать устойчивость к одному или больше антибиотиков, часто находятся в сложных транспозонах.Известные примеры сложных транспозонов Tn 5 и Тн 10 , определяющие устойчивость к канамицину и тетрациклин соответственно. Сложные транспозоны, вероятно, эволюционируют транспозиция гомологичных последовательностей вставок в близлежащие сайты в ДНК молекула.
Рисунок 5-11
Характеристики репрезентативных транспозонов (жирные линии) интегрированы в бактериальную хромосому (тонкие морщинки). Транспозоны являются важными генетическими элементами, потому что они вызывают мутации, опосредуют геномные перестройки, функционируют как переносимые области генетического (подробнее…)
Второй класс транспозонов состоит из высокогомологичных ТН Семейство . Эти транспозоны имеют более длинные (от 35 до 40 пара оснований) концевые инвертированные повторы, чем описанные сложные транспозоны выше, но в них отсутствуют концевые инсерционные последовательности. Все члены семьи кодируют функции транспозазы и резольвазы. Хорошо известные примеры из Tn Семейство транспозонов включает ампициллин резистентность транспозона Tn 3 и Tn 1000 (гамма-дельта транспозон), обнаруженный в F плазмида.Семья TnA занимает важное место в истории медицины. микробиология. Развитие высокой устойчивости к ампициллину у Haemophilus influenzae и Neisseria gonorrhoeae в 1970-е годы, что сильно ограничивало полезность ампициллина для лечения гонореи и гемофильной инфекции в регионах там, где такие штаммы стали преобладать, было вызвано распространением ампициллина детерминанты устойчивости из транспозонов Tn A в плазмидах Enterobacteriaceae к плазмидам в Haemophilus и Нейссерия .
Третий класс транспозонов состоит из бактериофага Mu и родственных ему умеренных фаги. Весь геном фага функционирует как транспозон, и репликация ДНК фага во время вегетативного роста происходит путем репликативной транспозиции. Интеграция профагов может происходить во многих разных участках бактериального хромосома и часто вызывает мутации. По этой причине Mu и родственные фаги иногда их называют фагами-мутаторами.
Четвертый класс транспозонов, обнаруженный у грамположительных бактерий и представлен Тн 917 , состоит из конъюгативных транспозонов которые полностью отличаются от транспозонов, описанных выше.В конъюгативный транспозон не создает дупликации целевой последовательности в которую он вставляется, а у грамположительных бактерий штамм-хозяин, несущий транспозон может действовать как супружеский донор. Бактерии-реципиенты не нужно близкородственен донорской бактерии. Транспозон иссекают из хромосома донора и передается путем конъюгации реципиенту, где он случайным образом интегрируется в хромосому. Tn 917 кодирует устойчивость к тетрациклину, но другие более крупные конъюгативные транспозоны могут кодировать дополнительная устойчивость к антибиотикам.Конъюгативные транспозоны, по-видимому, являются основным причина распространения устойчивости к антибиотикам у грамположительных бактерий.
Некоторые роли транспозонов в эволюции бактерий проиллюстрированы рассмотрением кишечные грамотрицательные бактерии и строение их плазмид. Бактерии собранные в доантибиотическую эру, содержали много плазмид, но они обычно отсутствовали детерминанты устойчивости. Многие из плазмид R из текущего клинические изоляты принадлежат к тем же группам несовместимости, что и обнаруженные плазмиды ранее, но они также определяют устойчивость к нескольким антибиотикам.В близкие отношения между их репликонами убедительно доказывают, что многие текущие плазмиды R произошли от более старых плазмид путем приобретения устойчивости детерминанты. Некоторые из плазмид с множественной устойчивостью к антибиотикам имеют индивидуальные транспозоны с несколькими детерминантами устойчивости, другие имеют несколько транспозоны резистентности расположены на разных участках, а третьи содержат сложные гибридные транспозоны устойчивости, образованные путем интеграции одного транспозона в другой. Поэтапное получение детерминант сопротивления может привести к в некоторых случаях к образованию составных транспозонов, кодирующих несколько детерминанты устойчивости.Терапевтическое использование антибиотиков и их включение в корм для животных обеспечивает селективные преимущества для бактерий с плазмидами R, тогда как конъюгация, трансформация и трансфекция обеспечивают средства для распространение плазмид R внутри и между видами бактерий. После плазмида, несущая транспозон, вводится в нового бактериального хозяина, транспозон и его детерминанты могут прыгать в хромосому или коренные плазмиды нового хозяина. Следовательно, стабильность мобилизующей плазмиды в новый бактериальный хозяин не важен для сохранения генетических детерминант расположен на транспозоне.
Рекомбинационная ДНК и клонирование генов
Существует множество методов создания гибридных молекул ДНК in vitro (рекомбинантная ДНК) и охарактеризовать их. Такие методы включают выделение определенных генов в гибридных репликоны, определяя их нуклеотидные последовательности и создавая мутации в обозначенные места (сайт-направленный мутагенез). Клон — это популяция организмы или молекулы, полученные в результате бесполого размножения от одного предка. Ген клонирование — это процесс включения чужеродных генов в репликоны гибридной ДНК.Клонированные гены могут быть экспрессированы в соответствующих клетках-хозяевах, и фенотипы, которые они определяют, что могут быть проанализированы. Некоторые ключевые концепции, лежащие в основе репрезентативных методов резюмируются здесь.
Первым шагом в клонировании генов является получение фрагментов донорской ДНК механическим или ферментативные методы. Некоторые эндонуклеазы рестрикции, обозначенные как класс II, являются особенно полезно для получения определенных фрагментов молекул ДНК. Они рассекаются обе нити двухцепочечных молекул ДНК в определенных палиндромных последовательностях (сайты рестрикции), длина которых обычно варьируется от четырех до восьми нуклеотидов, и полученные фрагменты ДНК называются рестрикционными фрагментами.Некоторое ограничение эндонуклеазы расщепляют совпадающие сайты с образованием фрагментов ДНК с тупыми концами, и другие разрезают в шахматном порядке для создания фрагментов ДНК с короткими, самокомплементарные одноцепочечные 5 ‘или 3’ концы (см.). Случайная вероятность того, что n соседние нуклеотиды в цепи ДНК будут соответствовать определенному сайту рестрикции составляет примерно 1/4 n . Сайты для ферментов, распознающих уникальные 4, 6 или 8 нуклеотидные мишени могут встречаться примерно один раз из каждых 256, 4096 или 65 536 нуклеотиды соответственно.Выбирая подходящие рестрикционные ферменты, специфическая ДНК молекулы, включая бактериальные хромосомы, плазмиды и геномы фагов, могут быть переваривается в наборы ограничивающих фрагментов, которые имеют соответствующие размеры для конкретных Приложения.
Таблица 5-3
Особенности ограничения типичного класса II Эндонуклеазы.
Карта ограничений определяет позиции целевых сайтов для конкретного ограничения эндонуклеазы в молекуле ДНК. Карты ограничений доступны для многих клонированных ДНК. фрагментов, плазмид и геномов фагов, а также для всей хромосомы E.coli и несколько других бактерий.
Второй этап клонирования генов — создание гибридных репликонов, состоящих из донорской ДНК. фрагменты и вектор клонирования (). Клонирующие векторы представляют собой небольшие репликоны плазмид или фагов, которые имеют один или несколько сайты рестрикции, в которые может быть вставлена чужеродная ДНК. Гибридные репликоны производится с использованием ДНК-лигазы для соединения рестриктированной векторной ДНК с донорной ДНК фрагменты, которые имеют совместимые концы, или, альтернативно, синтетические олигонуклеотиды используются в качестве линкеров для создания совместимости между донорскими и векторными молекулами ДНК с разными концами.Лигирование вектора с гетерогенным набором фрагментов ДНК из донорский геном называется клонированием дробовика, а сбор рекомбинантной ДНК молекулы, содержащие различные фрагменты, называются геномной библиотекой. Если специфический фрагмент ДНК доступен, он может быть включен в рекомбинантный репликон путем прямого клонирования в соответствующий вектор, выбранный из широкого разнообразия доступных векторов. Плазмидные и фаговые векторы используются в основном для клонирования небольших вставляет обычно меньше 10 кб. Примеры других векторов специального назначения включают космиды, которые представляют собой плазмидные векторы, которые могут быть упакованы в фаговые капсиды (лямбда космиды принимают вставки размером до 30-40 т.п.н.) и фагмиды, которые являются плазмид-фагами. гибридные репликоны, которые могут существовать в виде плазмид или одноцепочечных ДНК-фагов в разных условиях эксперимента.Космиды фага P1 могут принимать вставки до 100 т.п.н., а молекулы ДНК еще большего размера можно клонировать в дрожжевые искусственные хромосомы (YAC), которые могут стабильно поддерживать вставки размером до и более 1 Мбит / с в размер. Другие специализированные векторы обнаруживают промоторы, сигналы терминации транскрипции, или другие регуляторные элементы во вставках чужеродной ДНК или, наоборот, обеспечивают промоторы, с которых может быть инициирована транскрипция клонированных генов.
Рисунок 5-12
Схематическое изображение эксперимента по клонированию гена.Плазмидный вектор клонирования pBR322 имеет размер 4,36 пар оснований, имеет гены на устойчивость к ампициллину (amp r ) и тетрациклину ( tet r ) и имеет только один HindIII сайт рестрикции, расположенный внутри (подробнее …)
Заключительными этапами клонирования генов является введение гибридных репликонов в соответствующие клетки-реципиенты и проверить их на экспрессию интересующих донорских генов. Прокариотический клетки (включая бактерии) или эукариотические клетки (включая дрожжи, животные или растения) ячеек) могут использоваться в качестве получателей, но они различаются по своему вседозволенность для определенных репликонов, транскрипционные сигналы, которые они распознают и посттрансляционные модификации белковой структуры, которые они может выполнить.Можно вводить рекомбинантные молекулы ДНК, полученные in vitro. непосредственно в клетки-реципиенты путем трансформации или трансфекции. Кроме того, клоны в космидных или фаговых векторах могут быть упакованы в фаговые оболочки и введены в восприимчивые клетки-реципиенты путем трансдукции. Используя специализированные векторы (челнок векторов), которые могут реплицироваться в нескольких типах клеток, гены любого организма могут быть клонированы и обработаны в удобной бактериальной системе, а затем повторно введены в клетки исходного организма для анализа в их естественных окружающая обстановка.
Существует множество методов идентификации бактерий, содержащих рекомбинантную ДНК. молекулы. Большинство векторов клонирования имеют гены признаков, которые могут быть положительно выбран, например, устойчивость к антибиотикам. Кроме того, часто можно ввести чужеродную ДНК в вектор клонирования в сайт, который инактивирует несущественная, но легко узнаваемая векторная функция. Если оба эти условия выполняются, бактерии, содержащие рекомбинантные молекулы, могут быть отобраны и легко отличить от бактерий, содержащих только вектор.Бактерии в геномная библиотека, содержащая конкретный клонированный ген, может быть идентифицирована с помощью биохимические или иммунологические методы тестирования желаемого генного продукта. В качестве альтернативы интересующий клонированный ген может быть обнаружен непосредственно с помощью нуклеиновой кислоты. методы кислотной гибридизации при условии, что доступен конкретный ДНК- или РНК-зонд. Поскольку вставка чужеродной ДНК в вектор клонирования в соответствующем сайте делает не инактивировать его способность реплицироваться в соответствующих клетках-реципиентах, гибрид представляющие интерес репликоны могут быть амплифицированы путем репликации, и рекомбинантная ДНК молекулы или их генные продукты могут быть очищены и изучены.Способность очищать специфические молекулы ДНК сделали возможным развитие ферментативных и химических методов для определения их нуклеотидных последовательностей и современные методы введения мутации в определенных сайтах в клонированных генах основаны на знании их ограничения карты или нуклеотидные последовательности.
Методы рекомбинантной ДНК позволяют клонировать определенные фрагменты ДНК из любых источника в векторы, которые могут быть изучены на хорошо охарактеризованных бактериях, в эукариотические клетки, или in vitro .Применения клонирования ДНК быстро расширяется во всех областях биологии и медицины. В медицинской генетике такие область применения варьируется от пренатальной диагностики наследственных заболеваний человека до характеристика онкогенов и их роль в канцерогенезе. Фармацевтическая приложения включают крупномасштабное производство из клонированных человеческих генов биологических продукты с терапевтической ценностью, такие как полипептидные гормоны, интерлейкины и ферменты. Приложения в общественном здравоохранении и лабораторной медицине включают разработки вакцин для предотвращения конкретных инфекций и зондов для диагностики конкретных инфекции путем гибридизации нуклеиновых кислот или полимеразной цепной реакции (ПЦР).В в последнем процессе используются олигонуклеотидные праймеры и ДНК-полимераза для амплификации специфических последовательности ДНК-мишени во время нескольких циклов синтеза in vitro, что делает его возможно обнаружение редких целевых последовательностей ДНК в клинических образцах с большим чувствительность.
Регуляция экспрессии генов
Фенотипические свойства бактерий определяются их генотипами и ростом условия. Для бактерий в чистой культуре часто возникают изменения условий роста. в предсказуемых физиологических адаптациях у всех членов популяции.Как правило, основные генные продукты производятся в количествах, обеспечивающих самый быстрый рост данной среде, и продукты, необходимые при особых обстоятельствах, производятся только тогда, когда они нужны.
Физиологические адаптации часто связаны с изменениями метаболической активности. Поток метаболитов через определенные биохимические пути можно контролировать. как путем регулирования синтеза определенных ферментов, так и путем изменения активности существующих ферментов. Механизмы, регулирующие экспрессию генов, воздействуя на здесь обсуждается синтез конкретных генных продуктов.
Специфическая регуляция включает ген или группу генов, участвующих в конкретном метаболический процесс. Индукция и репрессия позволяют бактериям регулировать производство конкретных генных продуктов в ответ на соответствующие сигналы. Обычно катаболический ферменты индуцируются, когда субстрат для пути роста присутствует в ростовой среда, и биосинтетические ферменты подавляются продуктом пути. Ферменты, которые участвуют в одном биохимическом пути, часто занимают соседние положения на бактериальной хромосоме и координированно индуцируются или подавляются.Они образуют оперон, группу смежных генов, которые транскрибируются как единое целое. единица и переводится для получения соответствующих генных продуктов. Организация в оперон — важная стратегия для скоординированной регуляции экспрессии генов в бактериях. Опероны, которые могут быть индуцированы или подавлены, контролируются связыванием специфические регуляторные белки к определенным нуклеотидным последовательностям, которые функционируют как регуляторные сайты внутри оперона. Сравнение аминокислотных последовательностей многих этих различных регуляторных белков показали, что они могут быть сгруппированы вместе в семейства регуляторов (например,г. семейство белков lysR), которые могли развиться от общих предковых генов. Члены семейства lysR включают регуляторы таких разнообразные явления, такие как метаболизм лизина, цистеина и метионина у E. coli и репрессия железа в V. cholerae .
Глобальная регуляция одновременно изменяет экспрессию группы генов и оперонов, вместе называемые регулонами, которые контролируются одним и тем же регуляторным сигналом. Глобальное регулирование определяет реакцию бактерий на основные питательные вещества, такие как углерод, азот или фосфат, реакции на стрессы, такие как повреждение ДНК или тепло шока, а также синтез патогенами специфических факторов вирулентности при росте в их животные-хозяева.Количество определенного белка в бактериальной клетке может варьироваться. от нуля до многих тысяч молекул. Этот широкий диапазон часто определяется комбинированное действие нескольких регуляторных механизмов, влияющих на экспрессию соответствующий структурный ген. Регулирование достигается путем определения того, как часто ген транскрибируется в функциональную мРНК, насколько эффективно транслируется мРНК в белок, как быстро разлагается мРНК, как быстро белковый продукт переворачивается, и может ли активность белкового продукта быть изменена аллостерические эффекты или ковалентные модификации.
мРНК как транскрипционные единицы
Экспрессия гена начинается с ДНК-зависимой РНК-полимеразы (РНК-полимеразы) катализирует транскрипцию специфической мРНК из одной цепи ДНК-матрицы. Связывание РНК-полимеразы с ДНК происходит в определенных сайтах, называемых промоторами, и транскрипция начинается рядом с промотором. Сильные промоутеры могут взаимодействовать эффективно с РНК-полимеразой и инициировать транскрипцию с высокой скоростью; слабый промоторы инициируют транскрипцию с медленной скоростью. В любом случае мРНК синтезируется от его 5′-конца к его 3′-концу на приблизительно с постоянной скоростью, пока РНК-полимераза не распознает другой специфический сайт назвал терминатором.Затем РНК-полимераза отделяется от матрицы и транскрипция мРНК завершена.
Отдельные молекулы мРНК могут кодировать один или несколько полипептидов. Транскрипция оперон продуцирует полицистронную мРНК, которая кодирует несколько полипептидов. Трансляция полицистронных мРНК приводит к координированному синтезу кодируемых полипептиды, но каждый полипептид синтезируется как отдельная молекула. А специфический сайт связывания рибосомы расположен прямо перед началом каждого кодирующая последовательность на молекуле мРНК.
Информационные РНК в бактериях быстро разлагаются со средним периодом полураспада несколько минут, в отличие от тРНК и рРНК, которые намного более стабильны. Хотя мРНК составляют около половины вновь синтезированной РНК, они представляют собой только небольшая часть общей РНК. Короткий период полужизни мРНК имеет важные последствия для экспрессии генов. Если синтез определенной мРНК предотвращается, продукция соответствующих полипептидов быстро снижается.
Контроль экспрессии генов осуществляется путем регулирования одного или нескольких этапов путь от матрицы ДНК к продукту активного гена.Одновременный регулирование на нескольких уровнях позволяет лучше контролировать экспрессию генов, чем было бы возможно с помощью единого регулирующего механизма. Самый распространенный способ регулировать экспрессию генов в бактериях — контролировать производство специфических мРНК. Поскольку скорость удлинения молекулы РНК приблизительно равна константа, основными факторами, контролирующими синтез мРНК, являются скорость инициации и вероятность того, что полный текст будет произведено.
Регуляция инициации транскрипции
Некоторые мРНК в бактериях синтезируются с постоянной скоростью, что приводит к конститутивная продукция кодируемых полипептидов.Количество конкретных мРНК и полипептиды, продуцируемые разными конститутивными генами, сильно различаются, однако и часто отражают различия в силе сторонников тех гены.
Транскрипция многих оперонов регулируется в ответ на изменение окружающей среды. условия. Промоторы определяют максимальную скорость инициации транскрипции. для таких оперонов, но регуляторные белки участвуют в контроле транскрипция. Нуклеотидные последовательности в оперонах, к которым относятся специфические регуляторные связывание белков называется регуляторными сайтами или операторами.Операторы и промоутеры расположены близко друг к другу внутри оперонов и могут иметь перекрывающиеся ДНК последовательности. Связывание регуляторных белков с операторами может увеличиваться (положительное регулирование) или уменьшение (отрицательное регулирование) частоты инициация транскрипции. Белки, которые функционируют как негативные регуляторы: обычно называемые репрессорами. Поскольку регуляторные белки могут диффундировать через цитоплазма, структурные гены регуляторных белков не обязательно должны быть связаны к целевым оперонам.
Способность определять присутствие или отсутствие определенных соединений и изменять темпы синтеза соответствующих генных продуктов имеют центральное значение для контроля экспрессия гена. Регуляторные белки предлагают одно решение этой проблемы связь стимул-реакция. Многие регуляторные белки бифункциональны и связываются не только к соответствующим операторам, но и к конкретным эффекторам, которые небольшие молекулы, такие как определенные сахара, аминокислоты и другие метаболиты. Кроме того, регуляторные белки являются аллостерическими, что означает, что они могут существовать в различные конформации, которые проявляют различную аффинность связывания для своих родственные операторы и эффекторы.Достаточная концентрация эффекторных благовоний образование регуляторного комплекса белок-эффектор, который имеет либо высокий, либо низкое сродство к оператору в каждом конкретном случае. В отрицательно регулируемом систем, эффектор функционирует как корепрессор, если регулирующий белок-эффекторный комплекс является активным репрессором, а эффекторный комплекс функционирует как индуктором и вызывает дерепрессию, если свободный регуляторный белок является активным репрессор. Напротив, в положительно регулируемых системах эффектор стимулирует экспрессия оперона, если регуляторный белок-эффекторный комплекс представляет собой положительный регулятор, а эффектор подавляет экспрессию оперона, если свободный регуляторный белок является положительным регулятором.
Лактозный (lac) оперон E. coli является примером индуцибельный, отрицательно регулируемый оперон (). Ген lacI кодирует репрессор, который связывается с оператором lac и предотвращает транскрипцию из Промотор lac . Структурный ген этого репрессора отдельно от lac-оперона, и репрессор синтезируется конститутивно при низкая ставка. Когда индуктор связывается с lac-репрессором, комплекс не может связываться с оператора и не может предотвратить связывание РНК-полимеразы с промотором.При других благоприятных условиях экспрессируется оперон lac , в результате синтеза β-галактозидазы, β-галактозида пермеаза и β-галактозид трансацетилаза. Модель lac оперон может быть индуцирован лактозой или структурно родственными соединениями, такими как изопропил-b-D-тиогалактозид (IPTG). IPTG называют безвозмездным индуктором потому что он индуцирует lac-оперон, но не является субстратом для β-галактозидаза. Отрицательная регуляция также встречается во многих биосинтетических оперонов в E.coli . В таких оперонах продукт биосинтетический путь действует как эффектор для негативных регуляторных система.
Рисунок 5-13
Регулирование оперона lac в E. coli . Структурные гены lacZ , lacY и код lacA для β-галактозидазы, β-галактозид пермеаза и β-галактозид трансацетилаза, соответственно. Физиологическая роль lacA заключается в неизвестный. Репрессор lac (подробнее…)
Оперон арабинозы ( ara ) в E. coli является как положительно и отрицательно регулируется. В присутствии арабинозы нормативный белок стимулирует транскрипцию оперона ara. В отсутствие арабинозы однако регуляторный белок репрессирует оперон ara .
Опероны часто управляются более чем одним механизмом. Когда E. coli выращивают в среде, содержащей глюкозу и альтернативный источник углерода, такой как лактоза или арабиноза, индукция lac или ara оперон и использование лактоза или арабиноза задерживаются до тех пор, пока не будет израсходована глюкоза.Этот явление называется диуксическим ростом. Неспособность вызвать оперон lac или ara в присутствии глюкозы является примером репрессии катаболита. Лак и ara опероны положительно регулируются циклический-3 ‘, 5’-аденозинмонофосфат (цАМФ) и белок активатора гена катаболита (CAP) (продукт гена crp). В Комплекс цАМФ-ЦАП взаимодействует с сайтами связывания ЦАП в регуляторных областях некоторые опероны, в том числе опероны lac и ara, и стимулирует транскрипцию от соответствующих промоутеров.Уровень внутриклеточного цАМФ у E. coli высокий во время роста в отсутствие глюкозы и низкий во время роста в присутствии глюкозы. Катаболиты репрессии должны, следовательно, к отсутствию активации цАМФ-зависимых оперонов, когда бактерии выращены в присутствии глюкозы или другого быстро метаболизируемого углерода источники.
Регуляция терминации транскрипции
Аттенуация — это механизм регуляции оперонов путем прекращения транскрипции мРНК преждевременно.Аттенуация часто встречается в биосинтетических оперонах, включая trp , гистидин ( his ), треонин ( th ), изолейцин-валин ( ilv ) и фенилаланиновые (phe) опероны. Оперон trp в E. coli контролируется как репрессией, так и ослаблением. в наличие избытка триптофана, инициация транскрипции с trp Промотор репрессирован. Кроме того, однако, те транскрипты, инициированные с промотора trp , являются обычно заканчивается раньше любого из структурных генов Расшифровано trp, оперонов.Концентрация внутриклеточный триптофан, необходимый для поддержания репрессии, превышает уровень, необходимый для затухание. Такой двойной контроль позволяет клетке точно настроить выражение оперон trp в ответ на снижение концентрации триптофан.
Вторичная структура мРНК играет важную роль в механизме затухание. Все мРНК имеют лидерную последовательность между началом транскрипции. сайт и начало кодирующей последовательности для первого структурного гена.Для аминокислотные биосинтетические опероны, которые подвергаются аттенуации, лидер мРНК Последовательность имеет две отличительные особенности. Он кодирует короткий пептид, содержащий аминокислота, продуцируемая регулируемым путем, и она может образовывать альтернативу, взаимно несовместимые, двухцепочечные структуры РНК, которые участвуют в нормативные события. Например, пептид, кодируемый trp Лидерная последовательность мРНК содержит два соседних остатка триптофана, а пептид кодируемая лидерной последовательностью мРНК , имеет серию из семи последовательные остатки гистидина.показывает оперон trp и иллюстрирует альтернативный вторичный структур в лидерной последовательности trp мРНК . Есть три возможные вторичные структуры для этого региона, называемые сайтом паузы (сегменты 1 + 2), антитерминатор (сегменты 2 + 3) и аттенюатор (сегменты 3 + 4). Сегмент 1 сайта паузы перекрывается кодировкой область для пептида trpL . Какие второстепенные конструкции формируется зависит от эффективности трансляции trpL пептид.Когда сегменты 1 и 2 расшифровываются, они немедленно отжигаются и вызвать временную остановку РНК-полимеразы. Последующее инициирование трансляция пептида trpL нарушает сайт паузы и позволяет РНК-полимеразе продолжать транскрипцию. Если присутствует триптофан, транскрипция сегментов 3 и 4 и формирование структуры аттенюатора происходит, когда рибосома блокирует сегмент 2, в результате чего РНК-полимераза прекратить транскрипцию. Однако при дефиците триптофана триптофанил-тРНК также недостаточен, и рибосома останавливается на кодонах триптофана в сегменте 1.Это позволяет сегменту 2 отжигаться с вновь синтезированным сегментом 3, чтобы сформировать антитерминатор, тем самым делая сегмент 3 недоступным для отжига с сегментом 4. Таким образом предотвращается образование аттенюатора, и РНК-полимераза транскрибирует весь оперон trp . Таким образом истощение триптофан (фактически поставка триптофанил-тРНК) связан с регуляцией транскрипции биосинтетического оперона триптофана.
Рисунок 5-14
Регулировка оперона trp в E.coli . Организация оперона trp показана на верх фигуры. Пять структурных генов trpE , trpD , trpC , trpB и trpA кодируют ферменты которые катализируют терминальную последовательность реакций в триптофане формирование. (подробнее …)
В E. coli транскрипция и трансляция функционально спаренный. Бессмысленные мутации, вызывающие преждевременное прекращение трансляции часто вызывают снижение транскрипции более дистальных генов в том же опероне.Это явление называется полярностью. Рибосомы обычно инициируют трансляцию рост молекулы мРНК до завершения транскрипции, и такая трансляция маскирует сайты, которые в противном случае привели бы к прекращению действия РНК-полимеразы транскрипция. Преждевременное прекращение трансляции бессмысленным кодоном отделяет рибосомы от мРНК и позволяет РНК-полимеразе взаимодействовать с немаскированными сайтами терминации транскрипции.
В некоторых биологических системах, включая фаг лямбда, антитерминация используется как положительный регуляторный механизм для контроля экспрессии генов.Незамедлительно после заражение E. coli лямбда, РНК-полимераза связывается с двумя промоторов в лямбда-ДНК и инициирует дивергентные первичные транскрипты, которые оканчиваются на определенных сайтах лямбда-генома. Белок, кодируемый одним из первичные транскрипты взаимодействуют с РНК-полимеразой и позволяют ей продолжать транскрипция через первичные сайты терминации, тем самым выражая второй набор лямбда-генов. Один из продуктов, закодированный вторичной расшифровкой блокирует терминацию другой мРНК и активирует экспрессию третьего набора гены.Следовательно, антитерминация играет ключевую роль в управлении каскадом экспрессия гена во время литического роста фага лямбда. Антитерминация также участвует в регуляции оперонов рРНК E. coli.
Регуляция трансляции
Сайт связывания рибосомы на мРНК комплементарен последовательности на 3′-конец 16S рРНК. Взаимодействие между этими последовательностями облегчает формирование комплекса инициации синтеза белка. Как степень гомология с 16S рРНК и расстояние сайта связывания рибосомы от кодон инициации влияет на эффективность инициации трансляции.Использование кодонов в мРНК также влияет на эффективность трансляции. Информационные РНК для белков, которые требуются в больших количествах, как правило, используют кодоны, которые транслируются наиболее многочисленные виды тРНК, и обратное тоже верно.
Трансляционный контроль важен для регуляции синтеза рибосомных белки. Производство рибосом связано с высокими метаболическими затратами для бактерий, и при высоких скоростях роста рибосомы могут составлять почти половину клетки. масса. Большинство рибосомных белков и рРНК собраны в рибосомы, и пул свободных субъединиц рибосом очень мал.Гены рибосом белки организованы в несколько оперонов. Некоторые из свободных рибосом белки напрямую ингибируют трансляцию полицистронных мРНК, которые кодируют их, тем самым обеспечивая баланс синтеза рибосомальных белков с требование к их утилизации.
Регулоны и белки, передающие сигнал
Реглон — это группа генов или оперонов, контролируемых общим регулятором. Там есть несколько преимуществ в том, чтобы поставить под контроль разные опероны одним и тем же регулятор.Это позволяет связать восприятие одиночного стимула с экспрессия большого количества генов, которые могут быть необходимы для соответствующего ответ, и это устраняет потребность в скоординированно регулируемых генах быть сцепленным на бактериальной хромосоме. Стимул, к которому регулон Ответы могут быть внутриклеточным компонентом или сигналом окружающей среды. Отдельные опероны также могут регулироваться несколькими различными механизмы и выражаются в условиях, отличных от тех, которые влияют на весь регулон.
Более 40 различных регулонов были идентифицированы в E. coli . Конкретные примеры регулонов, которые отвечают на внутриклеточные компоненты, включают: описанный ранее регулон цАМФ-CAP и регулоны, контролируемые строгий ответ и ответ SOS. Когда рибосомы сталкиваются с незаряженной тРНК молекул во время синтеза белка активируется строгий ответ и приводит к быстрому прекращению синтеза рРНК. Новый нуклеотид под названием гуанозин-3′-дифосфат-5′-дифосфат (ppGpp) накапливается во время аминокислотного голодания.PpGpp, продуцируемый холостыми рибосомами, кажется, быть посредником строгого ответа, но точный механизм, вызывающий ингибирование синтеза рРНК неизвестно. Ответ SOS связан с повреждение ДНК и включает индукцию более 20 генов, участвующих в нескольких Пути репарации ДНК. Продукт гена recA обнаруживает ингибирование ДНК синтез и инициирует события, ведущие к протеолитическому расщеплению и инактивации репрессора пути SOS, кодируемого lexA ген.
Некоторые регулоны индуцируются определенными раздражителями окружающей среды, например, питательными веществами. ограничение или осмотический стресс. Часто опероны более чем одного регулона могут быть индуцированный, а термин «стимулон» использовался для описания набора генов, так что индуцированный. Обычно бактерии воспринимают такие условия окружающей среды двумя способами. компонентные системы. Первый компонент — это трансмембранный белок с внеклеточные и внутриклеточные домены. Его внеклеточный домен обнаруживает стимул окружающей среды, и его цитоплазматический домен передает сигнал.В второй компонент — бифункциональный цитоплазматический белок. Имеет домен-получатель который взаимодействует с модулем передатчика первого компонента, а также с эффекторный домен, контролирующий экспрессию соответствующего регулона. В модули передатчика и приемника двухкомпонентных систем регулирования от большое количество регулонов генетически связаны и имеют гомологию аминокислот. Сигнально-детектирующий и эффекторный домены белков из разных Однако регулоны различаются и определяют детектируемый сигнал и опероны, которые активируются или подавляются в ответ на этот сигнал.
Глобальное регулирование играет важную роль в физиологии патогенных бактерий. Например, Vibrio cholerae и Bordetella. pertussis выражают многие из детерминант вирулентности в управление системами передачи сигналов, относящимися к двухкомпонентной системы, описанные выше. Экспрессия белков, необходимых для инвазивного Фенотип контролируется температурой в Shigella . Yersinia enterocolitica чувствует температуры и концентрации ионов кальция и связывает эти сигналы с экспрессия генов и клеточная локализация продуктов генов, которые подходит для внутриклеточной или внеклеточной среды.В тканях хозяина концентрация свободного железа крайне низка, и большинство болезнетворных бактерий имеют системы транспорта железа с высоким сродством, которые индуцируются при низком уровне железа условия. Синтез дифтерийного токсина C. diphtheriae , токсин Shiga by Shigella dysenteriae , экзотоксин А от Pseudomonas aeruginosa и другие специфические белков у многих патогенных бактерий индуцируется в условиях ограниченного железом рост. Эти примеры показывают, как факторы окружающей среды могут регулировать экспрессия генов вирулентности у патогенных бактерий.
Генетика — Медицинская микробиология — Книжная полка NCBI
Генетическая информация в микробах
Генетический материал бактерий и плазмид — это ДНК. Бактериальные вирусы (бактериофаги или фаги) имеют ДНК или РНК в качестве генетического материала. Два основных Функции генетического материала — репликация и экспрессия. Генетический материал должен точно реплицироваться, чтобы потомство унаследовало все специфические генетические детерминанты (генотип) родительского организма. Выражение конкретных генетических материал в определенном наборе условий роста определяет наблюдаемые характеристика (фенотип) организма.Бактерии имеют мало структурных или особенности развития, которые можно легко наблюдать, но у них есть широкий спектр биохимические возможности и модели восприимчивости к противомикробным агентам или бактериофаги. Эти последние характеристики часто выбираются как наследуемые. черты, подлежащие анализу в исследованиях бактериальной генетики.
Структура нуклеиновой кислоты
Нуклеиновые кислоты — это большие полимеры, состоящие из повторяющихся нуклеотидных единиц (). Каждый нуклеотид содержит один фосфатная группа, один пентозный или дезоксипентозный сахар и один пурин или пиримидин основание.В ДНК сахар — это D-2-дезоксирибоза; в РНК сахар — это D-рибоза. В ДНК пуриновые основания представляют собой аденин (A) и гуанин (G), а пиримидиновые основания представляют собой тимин (Т) и цитозин (С). В РНК урацил (U) заменяет тимин. Химически модифицированные пуриновые и пиримидиновые основания обнаружены у некоторых бактерий и бактериофаги. Повторяющаяся структура полинуклеотидов включает чередование остатки сахара и фосфата, с фосфодиэфирными связями, связывающими 3′-гидроксильная группа одного нуклеотидного сахара к 5′-гидроксилу группа соседнего нуклеотидного сахара.Эти асимметричные фосфодиэфирные связи определяют полярность полинуклеотидной цепи. Основание пурина или пиримидина связан у 1′-углеродного атома каждого остатка сахара и выступает из повторяющийся сахарно-фосфатный остов. Двухцепочечная ДНК является спиральной, а две нити спирали антипараллельны. Двойная спираль стабилизирована водородные связи между пуриновыми и пиримидиновыми основаниями на противоположных цепях. В каждое положение, A на одной нити пары двумя водородными связями с T на противоположная нить, или G пары тремя водородными связями с C.Две нити поэтому двойные спирали ДНК комплементарны. Из-за взаимодополняемости двухцепочечная ДНК содержит эквимолярные количества пуринов (A + G) и пиримидины (T + C), где A равно T, а G равно C, но моль доля G + C в ДНК широко варьирует у разных бактерий. Информация в нуклеиновых кислотах кодируется упорядоченной последовательностью нуклеотидов. вдоль полинуклеотидной цепи, а в двухцепочечной ДНК последовательность каждого strand определяет, какой должна быть последовательность комплементарной цепи.В степень гомологии последовательностей между ДНК из разных микроорганизмов является самый строгий критерий для определения того, насколько тесно они связаны.
Рисунок 5-1
Двойная спиральная структура ДНК. На схеме представлена структура ДНК в виде спирали. лестница. Остов каждой полинуклеотидной цепи (представлен как лента) состоит из чередующихся остатков фосфата и дезоксирибозы связаны фосфодиэфиром (подробнее …)
Репликация ДНК
Во время репликации бактериального генома каждая нить в двойной спирали ДНК служит шаблоном для синтеза новой комплементарной цепи.Каждая дочь Таким образом, двухцепочечная молекула ДНК содержит одну старую полинуклеотидную цепь и одну вновь синтезированная нить. Такой тип репликации ДНК называется полуконсервативный. Репликация хромосомной ДНК у бактерий начинается с конкретный хромосомный участок, называемый источником, и продолжается двунаправленно, пока процесс завершен (). Когда бактерии делятся бинарным делением после завершения репликации ДНК, реплицированные хромосомы делятся на каждую из дочерних клеток. В области происхождения специфически и временно связываются с клеточной мембраной после того, как репликация ДНК была инициирована, что привело к модели, согласно которой мембрана прикрепление управляет разделением дочерних хромосом (модель репликона).Эти характеристики репликации ДНК во время роста бактерий соответствуют требования генетического материала к точному воспроизведению и наследуется каждой дочерней клеткой во время деления клетки.
Рисунок 5-2
Авторадиография интактной реплицирующейся хромосомы E. coli . Бактерии были радиоактивно помечены тритием тимидином для приблизительно в двух поколениях и подвергались бережному лизированию. Бактериальная ДНК затем был исследован авторадиографией. Вставка показывает репликацию бактериальный (подробнее…)
Экспрессия гена
Генетическая информация, закодированная в ДНК, выражается путем синтеза специфических РНК и белки, и информация течет от ДНК к РНК к белку. ДНК-направленный синтез РНК называется транскрипцией. Поскольку пряди двухспиральные ДНК антипараллельны и комплементарны, только одна из двух цепей ДНК может служат в качестве матрицы для синтеза конкретной молекулы мРНК. Информационные РНК (мРНК) передают информацию от ДНК, и каждая мРНК в бактериях функционирует как матрица для синтеза одного или нескольких конкретных белков.Процесс по нуклеотидная последовательность молекулы мРНК определяет первичный амино кислотная последовательность белка называется трансляцией. Рибосомы, комплексы рибосомные РНК (рРНК) и несколько рибосомных белков, транслируют каждую мРНК в соответствующую полипептидную последовательность с помощью транспортных РНК (тРНК), синтез аминоацил тРНК, факторы инициации и факторы элонгации. Все эти компоненты аппарата для синтеза белка функционируют в производство множества различных белков.Ген — это последовательность ДНК, которая кодирует белок, рРНК или молекула тРНК (продукт гена).
Генетический код определяет, как нуклеотиды в мРНК определяют аминокислоты. в полипептиде. Потому что в мРНК всего 4 разных нуклеотида. (содержащие U, A, C и G), отдельные нуклеотиды не содержат достаточно информации однозначно указать все 20 аминокислот. В динуклеотидах 16 (4 × 4) возможны расположения четырех нуклеотидов, а в Возможны 64 компоновки тринуклеотидов (4 × 4 × 4).Таким образом, требуется минимум три нуклеотида для обеспечения хотя бы одного уникального последовательность, соответствующая каждой из 20 аминокислот. В «Универсальный» генетический код, используемый большинством организмов (), представляет собой тройной код, в котором 61 из 64 возможных тринуклеотида (кодона) кодируют определенные аминокислоты, и любой из три оставшихся кодона (UAG, UAA или UGA) приводят к прекращению перевод. Кодоны, обрывающие цепь, также называются бессмысленными кодонами. потому что они не определяют никаких аминокислот. Генетический код описывается как вырожденный, потому что несколько кодонов могут использоваться для одной аминокислоты, и как неперекрывающиеся, потому что соседние кодоны не имеют общих нуклеотидов.Исключения из «универсального» кода включают использование UGA в качестве кодон триптофана у некоторых видов Mycoplasma и у митохондриальная ДНК и несколько дополнительных кодоновых различий в митохондриальной ДНК из дрожжей , Drosophila и млекопитающих. Трансляция мРНК осуществляется обычно инициируется кодоном AUG для метионина, а соседние кодоны транслируется последовательно, когда мРНК читается с 5 ‘по 3’ направление. Соответствующая полипептидная цепь собирается, начиная с ее аминоконце и продвигаясь к его карбоксиконцу.Последовательность амино кислоты в полипептиде, следовательно, колинеарны с последовательностью нуклеотидов в мРНК и соответствующем гене. Специфические ферментативные реакции участвуют в синтезе ДНК, РНК и белка, выходят за рамки этого глава.
Экспрессия генетических детерминант у бактерий включает однонаправленный поток информации от ДНК к РНК к белку. В бактериофагах либо ДНК, либо РНК. может служить генетическим материалом. При заражении бактерий РНК бактериофаги, молекулы РНК служат матрицами для репликации РНК и мРНК.Исследования ретровирусной группы вирусов животных показывают, что ДНК молекулы могут быть синтезированы из матриц РНК ферментами, обозначенными как РНК-зависимые ДНК-полимеразы (обратные транскриптазы). Этот поворот обычное направление потока генетической информации от РНК к ДНК, а не от ДНК в РНК — важный механизм передачи информации от ретровирусов. закодироваться в ДНК и стать частью геномов животных клетки.
Организация генома
Молекулы ДНК, которые реплицируются как дискретные генетические единицы в бактериях, называются репликоны.У некоторых штаммов Escherichia coli хромосома является в клетке присутствует только репликон. У других бактериальных штаммов есть дополнительные репликоны, такие как плазмиды и бактериофаги.
Хромосомная ДНК
Бактериальные геномы варьируются по размеру от примерно 0,4 × 10 9 до 8,6 × 10 9 дальтон (Да), некоторые из самых маленьких являются обязательными паразиты ( Mycoplasma ) и самые крупные, принадлежащие к бактериям способны к сложной дифференцировке, например, Myxococcus .В количество ДНК в геноме определяет максимальное количество информации, которую он может кодировать. Большинство бактерий имеют гаплоидный геном, единственную хромосому, состоящую из кольцевой двухцепочечной молекулы ДНК. Однако линейные хромосомы имеют был обнаружен у грамположительных Borrelia и Streptomyces spp., Один линейный и один круговой хромосома присутствует в грамотрицательной бактерии Agrobacterium tumefaciens . Единая хромосома общего кишечного бактерия E.coli составляет 3 × 10 9 Да (4500 пары килобаз [kbp]) размером примерно от 2 до 3 процентов от сухой массы клетки. Геном E. coli является всего около 0,1% от размера генома человека, но этого достаточно для кодирования несколько тысяч полипептидов среднего размера (40 кДа или 360 аминокислот).
Хромосома кишечной палочки имеет контурную длину приблизительно 1,35 мм, что в несколько сотен раз длиннее бактериальной клетки, но ДНК суперспиральный и плотно упакованный в бактериальный нуклеоид.Необходимое время для репликации всей хромосомы составляет около 40 минут, что составляет примерно вдвое меньше кратчайшего времени деления этой бактерии. ДНК репликация должна инициироваться так часто, как клетки делятся, чтобы быстро при выращивании бактерий новый виток хромосомной репликации начинается раньше, чем раньше. раунд завершен. При высоких темпах роста может быть четыре хромосомы. реплицируются с образованием восьми во время деления клеток, что связано с завершение раунда хромосомной репликации.Таким образом, хромосома в быстрорастущие бактерии размножаются более чем в одной точке. Репликация хромосомной ДНК бактерий сложна и включает в себя множество различных белки.
Плазмиды
Плазмиды — это репликоны, которые сохраняются как дискретные, внехромосомные генетические элементы в бактериях. Обычно они намного меньше бактериального хромосома, варьирующаяся от менее 5 до более нескольких сотен кб, хотя плазмиды размером до 2 Mbp встречаются у некоторых бактерий. Плазмиды обычно кодируют черты, которые не важны для жизнеспособности бактерий, и воспроизводятся независимо от хромосомы.Большинство плазмид имеют суперспиральную, круговую, двухцепочечные молекулы ДНК, но также были продемонстрированы линейные плазмиды у Borrelia и Streptomyces . Близко связанный или идентичные плазмиды демонстрируют несовместимость; они не могут быть стабильными сохраняется в том же бактериальном хозяине. Классификация плазмид основана на несовместимости или использования конкретных ДНК-зондов в тестах гибридизации с идентифицировать нуклеотидные последовательности, характерные для конкретной плазмиды репликоны.Некоторые гибридные плазмиды содержат более одного репликона. плазмиды кодируют функции, которые способствуют передаче плазмиды от донора бактерии к другим бактериям-реципиентам, но неконъюгативные плазмиды — нет. Конъюгативные плазмиды, которые также способствуют переносу бактериальной хромосомы из бактерии-доноры для других бактерий-реципиентов называются плазмидами фертильности, и обсуждаются ниже. Среднее количество молекул данной плазмиды на бактериальная хромосома называется числом ее копий.Большие плазмиды (> 40 пары килобаз) часто бывают конъюгативными, имеют уменьшенное количество копий (от 1 до нескольких на хромосома), код для всех функций, необходимых для их репликации, и разделяются между дочерними клетками во время деления клеток способом похож на бактериальную хромосому. Плазмиды размером менее 7,5 тысяч пар оснований обычно не сопряжены, имеют большое количество копий (обычно 10–20 на хромосому), полагаются на своего бактериального хозяина, чтобы обеспечить некоторые необходимые функции для репликации и случайным образом распределяются между дочерними клетками на разделение.
Многие плазмиды контролируют важные с медицинской точки зрения свойства патогенных бактерий, включая устойчивость к одному или нескольким антибиотикам, выработку токсинов и синтез структур клеточной поверхности, необходимых для прикрепления или колонизации. Плазмиды, определяющие устойчивость к антибиотикам, часто называют плазмидами R. (или R-факторы). Типичные токсины, кодируемые плазмидами, включают термолабильные и термостойкие энтеротоксины E. coli , эксфолиативный токсин золотистый стафилококк и столбнячный токсин Clostridium tetani .Некоторые плазмиды загадочны и не имеют узнаваемое воздействие на бактериальные клетки, в которых они находятся. Сравнение плазмиды профили — полезный метод оценки возможного родства отдельных клинические изоляты определенного вида бактерий для эпидемиологических исследований. Обсуждается роль плазмид в развитии устойчивости к антибиотикам. ниже.
Бактериофаги
Бактериофаги (бактериальные вирусы, фаги) — инфекционные агенты, которые размножаются как облигатные внутриклеточные паразиты у бактерий.Внеклеточные фаговые частицы метаболически инертны и состоят в основном из белков и нуклеиновой кислоты (ДНК или РНК, но не то и другое вместе). Белки фаговой частицы образуют защитную оболочка (капсид), окружающая плотно упакованный геном нуклеиновой кислоты. Фаг геномы различаются по размеру от примерно 2 до 200 килобаз на цепь нуклеиновой кислоты. кислоты и состоят из двухцепочечной ДНК, одноцепочечной ДНК или РНК. Фаг геномы, как и плазмиды, кодируют функции, необходимые для репликации в бактериях, но в отличие от плазмид они также кодируют белки капсида и неструктурные белки требуется для сборки фага.Несколько морфологически различных типов фага были описаны, в том числе многогранные, нитевидные и сложные. Сложный фаги имеют многогранные головы, к которым хвосты, а иногда и другие придатки (хвост пластины, хвостовые волокна и т. д.).
Один цикл роста фага показан на. Инфекция инициируется адсорбцией фага на специфических рецепторы на поверхности восприимчивых бактерий-хозяев. Капсиды остаются на поверхность клетки, а геномы ДНК или РНК проникают в клетки-мишени (проникновение).Поскольку инфекционность геномной ДНК или РНК намного меньше, чем у зрелых вируса, сразу после заражения наступает период, называемый периодом затмения. во время которого невозможно обнаружить внутриклеточный инфекционный фаг. Заражение РНК или ДНК фага реплицируются, чтобы произвести множество новых копий генома фага, и продуцируются фаг-специфические белки. Для большинства фагов сборки потомства происходит в цитоплазме, а высвобождение потомства происходит путем лизиса клеток. В напротив, нитчатые фаги образуются на клеточной оболочке и высвобождаются не убивая клетки-хозяева.Период затмения заканчивается, когда внутриклеточный появляются заразные потомства. Скрытый период — это интервал от заражения. до появления внеклеточного потомства, а период роста — это интервал от конец латентного периода, пока все фаги не станут внеклеточными. Среднее число частиц фага, продуцируемых каждой инфицированной клеткой, называемого размером взрыва, составляет характерна для каждого вируса и часто составляет от 50 до нескольких сотен. Для обсуждения строения, размножения и классификации животных. вирусы, см. главы 41 и 42.
Рисунок 5-3
Одностадийный рост бактериофага. Культура восприимчивых бактерий синхронно заражается бактериофаг добавлен в момент времени 0 при низкой множественности инфекции. Вскоре после этого неабсорбированный фаг инактивируется добавлением антифаговая антисыворотка, (подробнее …)
Фаги подразделяются на две основные группы: вирулентные и умеренные. Рост вирулентные фаги у чувствительных бактерий разрушают клетки-хозяева. Заражение восприимчивые бактерии к умеренным фагам могут иметь один из двух результатов: литический рост или лизогения.Литический рост умеренных и вирулентных бактериофагов аналогично, что приводит к образованию фагового потомства и гибели бактерий-хозяев. Лизогения — это особый тип латентной вирусной инфекции, при которой геном фага реплицируется как профаг в бактериальной клетке. У большинства лизогенных бактерий гены, необходимые для развития литического фага, не экспрессируются, и производство инфекционный фаг не встречается. Кроме того, лизогенные клетки невосприимчивы к суперинфекция вирусом, который они носят как профаги.Физическое состояние профага не является идентичным для всех вирусов умеренного климата. Например, профаг бактериофага λ в E. coli интегрирован в бактериальную хромосому в определенном месте и реплицируется как часть бактериальная хромосома, тогда как профаг бактериофага P1 у E. coli реплицируется как внехромосомная плазмида.
Рост литического фага происходит спонтанно в небольшой фракции лизогенных клеток, и несколько внеклеточных фагов присутствуют в культурах лизогенных бактерий.Для некоторых лизогенных бактерий синхронная индукция развития литических фагов встречается во всей популяции лизогенных бактерий, когда они обрабатываются агенты, которые повреждают ДНК, такие как ультрафиолет или митомицин С. Потеря профаг из лизогенной бактерии, переводя его в нелизогенное состояние и восстановление восприимчивости к инфекции первоначально присутствующим фагом как профаг, называется лечением.
Некоторые фаги умеренного пояса содержат гены бактериальных характеристик, которые не связаны с развитием литического фага или лизогенным состоянием, и экспрессия такие гены называют фаговой конверсией (или лизогенной конверсией).Примеры конверсия фагов, которая важна для микробной вирулентности, включает производство токсина дифтерии Corynebacterium diphtheriae , эритрогенный токсин Streptococcus pyogenes (группа А β-гемолитические стрептококки), ботулинический токсин от Clostridium botulinum , а шига-подобные токсины — E. coli . В В каждом из этих примеров ген, кодирующий бактериальный токсин, присутствует в умеренный геном фага. Специфичность O-антигенов в Salmonella также можно контролировать с помощью фаговой конверсии.Типирование фагов — это тестирование штаммов определенного вида бактерий на восприимчивость к специфическим бактериофагам. Паттерны восприимчивости к набор типирующих фагов предоставляет информацию о возможном родстве отдельные клинические изоляты. Такая информация особенно полезна для эпидемиологические исследования.
Мутация и отбор
Мутации — это наследуемые изменения генома. Спонтанные мутации у человека бактерии встречаются редко. Некоторые мутации вызывают изменения фенотипических характеристик; то о возникновении таких мутаций можно судить по вызываемым ими эффектам.В контрольные организмы, специфичные для микробной генетики, обозначены как штаммы дикого типа, а потомки, в геномах которых есть мутации, называются мутантами. Таким образом, мутанты характеризуются унаследованными различиями между ними и их предковые штаммы дикого типа. Вариантными формами конкретной генетической детерминанты являются: называются аллелями. Генотипические символы — строчные буквы, сокращения выделены курсивом. указать отдельные гены с надстрочным индексом (+), указывающим на дикий тип аллель. Фенотипические символы пишутся с большой буквы, а не курсивом, чтобы их можно было различить. от генотипических символов.Например, генотипический символ способности производить β-галактозидаза, необходимая для ферментации лактозы, является lacZ + и мутанты, которые не могут производить β-галактозидазы lacZ . Фенотип ферментации лактозы обозначается Lac + , а неспособность ферментировать лактозу Lac — .
Обнаружение мутантных фенотипов
Селективные и дифференциальные среды полезны для выделения бактериальных мутантов. Некоторые селективные среды позволяют расти определенным мутантам, но не позволяют штаммы дикого типа для выращивания.Редкие мутанты можно выделить с помощью таких селективных СМИ. Дифференциальные среды позволяют бактериям дикого типа и мутантным бактериям расти и формироваться. колонии, различающиеся по внешнему виду. Обнаружение редких мутантов по дифференциалу media ограничено общим количеством колоний, которые можно наблюдать. Рассматривать штамм E. coli дикого типа, чувствительный к антибиотик стрептомицин (фенотип Str s ) и может использовать лактозу в качестве единственный источник углерода (фенотип Lac + ).Спонтанно Встречающиеся мутанты Str r редки и обычно встречаются с частотами менее одной на 109 бактерий в культурах дикого типа E. coli . Тем не менее, мутанты Strr можно легко выделить, используя селективная среда, содержащая стрептомицин, поскольку Str s дикого типа бактерии погибают. Выделение лактозонегативных (фенотип Lac — ) мутанты E. coli представляют собой другая проблема. На минимальных средах с лактозой в качестве единственного источника углерода, Lac + штаммы дикого типа будут расти, но Мутанты Lac — не могут расти.На дифференциальных средах, таких как МакКонки-лактозный агар или эозин-метиленовый синий-лактозный агар, Lac + дикого типа и мутантные штаммы Lac — E. coli можно отличить по цвету, но самопроизвольно Мутанты Lac — слишком редки, чтобы их легко выделить. Селективная среда для мутантов Lac — E. coli можно получить путем включения химических аналогов лактозы, которые превращаются в токсичные метаболиты бактериями Lac + , но не мутантами Lac —.Мутанты Lac — затем могут расти на такой среде, но бактерии дикого типа Lac + убиты.
Мутации, инактивирующие основные гены гаплоидных организмов, обычно летальные, но такие потенциально смертельные мутации часто можно изучить, если их экспрессия контролируется манипулированием экспериментальными условиями. Для Например, мутация, которая увеличивает термолабильность важного гена продукт может предотвратить рост бактерий при 42 ° C, хотя мутант бактерия все еще может расти при 25 ° C.И наоборот, чувствительные к холоду мутанты выражают мутантный фенотип при низкой температуре, но не при высокой температуре. Чувствительные к температуре и холоду мутации являются примерами условных мутации, как и подавляемые мутации, описанные далее в этой главе. А условный летальный фенотип указывает на то, что мутантный ген важен для жизнеспособность.
Спонтанные и индуцированные мутации
Скорость мутации у бактерий определяется точностью репликации ДНК, возникновение повреждения ДНК и эффективность механизмов восстановления поврежденной ДНК.
Для конкретного бактериального штамма в определенных условиях роста мутация скорость для любого конкретного гена постоянна и выражается как вероятность мутации на деление клетки. В популяции бактерий, выросших из небольшого inoculum, доля мутантов обычно прогрессивно увеличивается по мере увеличения размера бактериальной популяции увеличивается.
Мутации у бактерий могут происходить спонтанно и независимо от для их обнаружения используются экспериментальные методы. Этот принцип был впервые продемонстрирован тестом на колебания ().В количество устойчивых к фагам мутантов E. coli в повторностях культуры, выращенные из небольших инокул, были измерены и сравнены с культурами, выращенными в несколько образцов, взятых из одной культуры. Если мутации по устойчивости к фагам произошла только после воздействия фага, вариабельность количества мутантов между культурами должны быть одинаковыми в обоих наборах условий. Напротив, если устойчивые к фагам мутанты возникли спонтанно до воздействия от бактерий к фагу, количество мутантов должно быть более вариабельным в самостоятельно выращенные культуры, поскольку различия в размерах бактерий популяции, когда появился первый мутант, будет способствовать наблюдаемому изменчивость.Данные показали, что мутации устойчивости к фагам в E. coli возник спонтанно с постоянной вероятностью на деление клетки.
Рисунок 5-4
Испытание на колебания. Различия в количестве колоний устойчивых к фагам мутантов в повторные образцы из одной субкультуры были небольшими и отражали только ожидаемые колебания из-за ошибок выборки. Напротив, количество устойчивых к фагам колоний (подробнее …)
Посев на реплики подтвердил, что мутации у бактерий могут возникать спонтанно, без воздействия на бактерии селективных агентов ().Для покрытия реплик: плоский стерильный вельветовый поверхность используется для сбора инокулята с поверхности эталонной чашки с агаром и перенесите образцы в другие чашки с агаром. Таким образом, образцы Популяция бактерий с мастер-планшета переносится на реплики не искажая их пространственного расположения. Если реплики содержат селективной среды и мастер-планшетов нет, положения отобранных мутантов можно отметить колонии на чашках-репликах и бактерии, которые не подвергались воздействию к условиям отбора могут быть изолированы с тех же позиций на мастер пластина.Мутанты E. coli , устойчивые к бактериофагу Т1 или стрептомицину были выделены таким образом, не подвергая бактерии дикого типа к бактериофагу или антибиотику.
Рисунок 5-5
Выявление ранее существовавших бактериальных мутантов по реплике покрытие. Мастер-планшет был сильно засеян образцом из чистых культур чувствительные к фагам бактерии. После инкубации бактерии от хозяина планшеты были перенесены путем посева реплик в дубликаты (подробнее …)
На частоту мутаций влияют как экологические, так и генетические факторы.Контакт с бактерий в мутагенные агенты вызывает увеличение скорости мутаций, иногда на на несколько порядков. Многие химические и физические агенты, включая рентгеновские лучи и ультрафиолет, обладают мутагенной активностью. Канцерогенные химические вещества для животных часто являются мутагенными для бактерий или могут быть преобразованы животными ткани на метаболиты, которые являются мутагенными для бактерий. Стандартизированные тесты для мутагенность бактерий используются в качестве скрининговых процедур для выявления агенты окружающей среды, которые могут быть канцерогенными для человека.Гены-мутаторы в бактерии вызывают увеличение частоты спонтанных мутаций для самых разных другие гены. Экспрессия этих генов, вызванная повреждением ДНК (см. SOS-ответ позже), позволяет восстанавливать повреждения ДНК, которые в противном случае были бы летальными, но за счет подверженный ошибкам механизм, который увеличивает скорость мутации. В целом скорость мутации — вероятность того, что мутация произойдет где-то в бактериальный геном на деление клетки — относительно постоянен для множество организмов с геномами разного размера и, по-видимому, важный фактор в определении пригодности бактериального штамма к выживанию в природе.Большинство мутаций вредны, и существует риск неблагоприятных мутаций для отдельные бактерии должны быть сбалансированы с положительным значением изменчивости, поскольку механизм адаптации бактериальных популяций к изменяющимся условиям окружающей среды. условия.
Молекулярная основа мутаций
Мутации классифицируются на основе структурных изменений, происходящих в ДНК (). Некоторые мутации локализованы в коротких сегментах ДНК (например, при нуклеотидных заменах, микроделеции и микровставки).Другие мутации затрагивают большие области ДНК и включают делеции, вставки или перестройки сегментов ДНК.
Когда нуклеотидная замена происходит в области ДНК, которая кодирует полипептид, один из трех нуклеотидов в одном кодоне соответствующая молекула мРНК будет изменена. Тихие мутации не меняют структура или функция полипептида, потому что один кодон в мРНК заменен на другой для той же аминокислоты. Другие замены вызывают замену одной аминокислоты. заменен другим в конкретном положении в полипептиде соответствующий измененному кодону.Мутации, приводящие к замене одного аминокислота для другой в полипептидной цепи называется миссенс-мутациями. Влияние аминокислотных замен на функцию полипептидного гена продукт различается и зависит от местоположения и идентичности аминокислоты замена. Мутантные полипептиды, содержащие замены аминокислот, обычно имеют общие антигенные детерминанты с полипептидом дикого типа и часто имеют некоторые остаточная биологическая активность. Мутации, приводящие к замене аминокислоты кислотный кодон с терминирующим кодоном называют нонсенс-мутациями.Это результаты в продукции амино-концевого фрагмента нормального полипептида, когда мутантная мРНК транслируется. Бессмысленные мутации часто приводят к полной потере активность генного продукта.
Из-за триплетной природы генетического кода последствия мутаций вызванные вставками или удалениями небольшого количества нуклеотидов (микровставки, микроделеции) зависят как от количества, так и от последовательности вовлеченные нуклеотиды. Удаление или добавление кратных трех нуклеотидов пары не влияют на рамку считывания, но вызывают удаление или добавление соответствующее количество аминокислот в одном сайте полипептида.Если новый кодон обрыва цепи вводится, преждевременное обрыв цепи происходит в пределах полипептид. Напротив, добавление или удаление других номеров пары нуклеотидов изменяют рамку считывания для всего сегмента мРНК от мутация дистального конца гена. Следовательно, мутации сдвига рамки считывания может вызвать резкие изменения в структуре и активности полипептида генные продукты, и их часто классифицируют как бессмысленные мутации.
Тесты комплементации
Чтобы определить, находятся ли мутации в одном и том же гене или в разных генах, тесты комплементации проводятся с частично диплоидными бактериальными штаммами ().Две копии региона бактериальной хромосомы, несущей мутацию, присутствуют в том же бактерия, причем каждая копия содержит разные мутации (мутации находятся в транс расположение). Фенотип дикого типа указывает на то, что мутации разные гены. Это явление называется комплементацией. Если мутант фенотипа, необходимо провести контрольный эксперимент с мутации в цис-расположении, чтобы исключить возможность того, что дикий тип аллели не могут нормально экспрессироваться в частично диплоидном штамме бактерий.Изначально тесты комплементации назывались «цис-транс». тесты, а термин цистрон иногда используется как синоним гена. Дополнительные тесты могут быть выполнены и интерпретированы, даже если биохимические функции продуктов гена неизвестны.
Рисунок 5-6
Комплементация — это метод проверки функционального гена товары. Два мутанта со схожими фенотипами (неспособность конвертировать субстрат X к продукту Z). Мутации в этих штаммах обозначены a и b, соответственно, и аллели дикого типа (подробнее…)
В качестве примера рассмотрим использование теста дополнения для характеристики двух независимо полученные мутанты Lac — E. coli . Биохимический путь утилизации лактозы требует β-галактозид пермеаза (генотипический символ lac Y) для транспортировки лактоза в бактериальную клетку и β-галактозидазу (генотипический символ lacZ ) для превращения лактозы в D-глюкозу и D-галактозу. Мутанты без β-галактозид пермеазы или β-галактозидазы не могут используйте лактозу для роста.Если мутации в обоих Lac — мутанты инактивировали тот же белок (например, β-галактозид), затем частичный диплоидный штамм, содержащий генов lacZ от обоих мутанты в транс-структуре не могут использовать лактозу. В напротив, если бы генотипы двух мутантов были lacZ + lacY и lacZ lacY + , частично диплоидная бактерия будет производить активную β-галактозидазу из lacZ + определитель и активный β-галактозид пермеаза из лаковый + определитель.Дополнение произойдет, и частично диплоидный штамм будет использовать лактозу.
Реверсия и подавление
Мутации, которые преобразуют фенотип из дикого типа в мутантный, называются прямыми мутации и мутации, которые изменяют фенотип с мутантного обратно на дикий тип называются обратными мутациями (реверсиями). Бактериальные штаммы, содержащие обратные мутации называются ревертантами. Анализ мутаций, вызывающих фенотипическая реверсия дает полезную информацию. Обратные мутации, восстанавливающие точные нуклеотидные последовательности ДНК дикого типа являются истинными реверсиями.Истинный ревертанты идентичны штаммам дикого типа как генотипически, так и фенотипически. Обратные мутации, которые не восстанавливают точный нуклеотид Последовательности ДНК дикого типа называются супрессорными мутациями (супрессорами). Некоторые ревертанты, несущие супрессорные мутации, фенотипически неотличимы от штаммов дикого типа. Другие ревертанты, называемые псевдоревертанты, фенотипически отличимые от штаммов дикого типа, например, из-за тонких различий в характеристиках ферментативного восстановленная активность (например, удельная активность, субстрат специфичность, кинетические константы или восприимчивость к термическому или химическому инактивация).Распознавание псевдоревертантных фенотипов предполагает наличие супрессорных мутаций.
Супрессорные мутации могут быть внутригенными или экстрагенными. Внутригенные супрессоры расположены в том же гене, что и подавляемые ими прямые мутации. В возможные места и характер внутригенных супрессоров определяются исходная прямая мутация и отношения между первичной структурой продукта гена и его биологической активности. Экстрагенные супрессоры находятся в разных генах от мутаций, эффекты которых они подавляют.В способность экстрагенных супрессоров подавлять множество независимых мутаций можно протестировать. Некоторые экстрагенные супрессоры специфичны для определенных генов, некоторые специфичны для определенных кодонов, а некоторые имеют другую специфичность узоры. Экстрагенные супрессоры, которые меняют фенотипические эффекты кодоны, завершающие цепь, были хорошо охарактеризованы, и было обнаружено, что они изменяют структура специфических тРНК. Конкретная супрессорная тРНК может разрешать определенные кодон обрыва цепи, который будет транслироваться, что приведет к включению специфической аминокислоты в формирующийся полипептид в положении, соответствующем к кодону обрыва цепи.У бактерии, имеющей оканчивающуюся цепь мутация и соответствующий экстрагенный супрессор, трансляция мРНК содержащий мутантный кодон, следовательно, может привести к образованию полноразмерного полипептид. Биологическая активность полноразмерного полипептида, образованного как Последствия подавления зависят как от количества вырабатываемого белка, так и от функциональные последствия замены конкретной аминокислоты, определяемые супрессорная тРНК.
Обмен генетической информацией
Биологическое значение сексуальности у микроорганизмов заключается в увеличении вероятность того, что редкие независимые мутации произойдут вместе в одном микробе и подвергнуться естественному отбору.Генетические взаимодействия между микробами позволяют их геномы эволюционируют гораздо быстрее, чем за счет одной лишь мутации. Представитель явления, имеющие медицинское значение, которые включают обмен генетической информацией или геномные перестройки включают быстрое появление и распространение антибиотика. плазмиды устойчивости, вариации жгутиковых фаз у сальмонелл и антигенные изменение поверхностных антигенов у Neisseria и Borrelia.
Половые процессы у бактерий включают передачу генетической информации от донора к реципиент и приводит либо к замене аллелей донора на аллели реципиента или добавление донорских генетических элементов в геном реципиента.Трансформация, трансдукция и конъюгация — это половые процессы, в которых используются разные механизмы для ввести донорскую ДНК в реципиентные бактерии (). Поскольку донорская ДНК не может сохраняться в бактерии-реципиенте, если это часть репликона, рекомбинация между геномами донора и реципиента часто требуется для получения стабильного гибридного потомства. Наиболее вероятна рекомбинация когда донорская и реципиентная бактерии происходят от одного или близкородственного происхождения разновидность.
Рисунок 5-7
Обмен генетической информацией в бактериях.Трансформация, трансдукция и конъюгация различаются способами введение ДНК из клетки-донора в клетку-реципиент. А) В трансформация, отбираются фрагменты ДНК, освобожденные от донорских бактерий вверх по компетентному (подробнее …)
Для того, чтобы рекомбинант был обнаружен, его фенотип должен отличаться от обоих родительских фенотипы. Перед рекомбинантным фенотипом может потребоваться рост или деление клеток. выражается. Задержка экспрессии рекомбинантного фенотипа до гаплоида сегрегация рекомбинантного генома называется задержкой сегрегации и задержкой до тех пор, пока синтез продуктов, кодируемых донорскими генами, называется фенотипическим отставанием.Тестирование на сцепление (неслучайная перегруппировка родительских аллелей в рекомбинантных потомство) возможно, когда родительские бактерии имеют разные аллели для нескольких гены. Донорский аллель невыбранного гена с большей вероятностью присутствует в рекомбинантов, если он связан с выбранным донорским геном, чем если он не связан с выбранный донорский ген. Количественный анализ связей позволяет построить генетические карты. Геном E. coli является кольцевым (), что определяется как генетическими сцепление и прямой биохимический анализ хромосомной ДНК, и генетическая карта колинеарно с физической картой хромосомной ДНК.Генетическое и физическое картирование также используются для анализа внехромосомных репликонов, таких как бактериофаги и плазмиды.
Рисунок 5-8
Круговая генетическая карта E. coli . Позиции репрезентативных генов указаны на внутреннем круге. Расстояния между генами калибруются в минутах, в зависимости от времени. требуется для переноса при конъюгации. Положение треонина (th) локус произвольно (подробнее …)
Многие бактерии имеют системы рестрикции-модификации (РМ), состоящие из модифицирующих ферменты, которые метилируют остатки аденина или цитозина в определенных последовательностях в их собственная ДНК и соответствующие эндонуклеазы рестрикции, которые расщепляют чужеродную ДНК, которая не несет специфической модификации в тех же целевых последовательностях.Некоторые рестрикционные ферменты будут расщеплять только ДНК, метилированную в определенных последовательности. Эти системы ограничения, которые могли развиться для защиты бактерий против вторжения фагов или плазмид, являются важным барьером для генетических обмены между различными бактериальными штаммами или видами. Недавние данные свидетельствуют о что плазмидные системы RM могут быть способом для плазмиды обеспечить ее перенос в штамм-хозяин, поскольку клетки, теряющие плазмиду (и соответствующие защитные ген метилазы) убиваются под действием более стабильного рестрикционного фермента, который атакует недавно реплицированную, но неизмененную хромосомную ДНК.
Трансформация
При трансформации захватываются фрагменты ДНК, высвобожденные из донорских бактерий. непосредственно из внеклеточной среды бактериями-реципиентами. Рекомбинация происходит между отдельными молекулами трансформирующей ДНК и хромосомами реципиентные бактерии. Чтобы быть активными в трансформации, молекулы ДНК должны находиться на длиной не менее 500 нуклеотидов, и трансформирующая активность быстро разрушается обработкой ДНК дезоксирибонуклеазой. Молекулы трансформирующей ДНК соответствуют к очень маленьким фрагментам бактериальной хромосомы.Котрансформация генов поэтому маловероятно, если они не связаны настолько тесно, что могут быть кодируется на одном фрагменте ДНК. Трансформация была обнаружена в Streptococcus pneumoniae и встречается у других бактериальных родов, в том числе Haemophilus , Neisseria , Bacillus и Staphylococcus . Способность бактерий для поглощения внеклеточной ДНК и трансформации, называемой компетентность зависит от физиологического состояния бактерий.Многие бактерии которые обычно некомпетентны, могут быть использованы для анализа ДНК в лаборатории манипуляции, такие как кальциевый шок или воздействие высоковольтного электрического пульс (электропорация). У некоторых бактерий (включая Haemophilus и Neisseria) Поглощение ДНК зависит от наличия определенных олигонуклеотидных последовательностей в трансформирует ДНК, но у других (включая Streptococcus pneumoniae) захват ДНК не зависит от последовательности. Компетентные бактерии также могут занимать неповрежденные ДНК бактериофага (трансфекция) или плазмидная ДНК, которая затем может реплицироваться как внехромосомные генетические элементы в бактериях-реципиентах.Напротив, фрагмент хромосомной ДНК от бактерии-донора обычно не может реплицироваться в бактерия-реципиент, если она не станет частью репликона путем рекомбинации. Исторически сложилось так, что характеристика «трансформирующего принципа» из S. pneumoniae при условии, что первое прямое доказательство наличия ДНК генетический материал.
Трансдукция
При трансдукции бактериофаги действуют как векторы для введения ДНК от донора бактерии в бактерии-реципиенты путем инфицирования. Для некоторых фагов, называемых генерализованные трансдуцирующие фаги, небольшая фракция вирионов, продуцируемых во время литический рост аберрантный и содержит случайный фрагмент бактериального генома вместо фаговой ДНК.Каждый индивидуальный трансдуцирующий фаг несет свой набор тесно связанных генов, представляющих небольшой сегмент бактериального генома. Трансдукция, опосредованная популяциями таких фагов, называется генерализованной. трансдукция, потому что каждая часть бактериального генома имеет приблизительно такая же вероятность передачи от донора к бактериям-реципиенту. Когда генерализованный трансдуцирующий фаг заражает реципиентную клетку, экспрессия передаются донорские гены. Абортивная трансдукция относится к временным экспрессия одного или нескольких донорских генов без образования рекомбинантного потомства, тогда как полная трансдукция характеризуется образованием стабильных рекомбинанты, которые наследуют донорские гены и сохраняют способность их экспрессировать.В прерванная трансдукция донорский фрагмент ДНК не реплицируется, и среди потомство исходного трансдуктанта только одна бактерия содержит донорскую ДНК фрагмент. У всех остальных потомков продукты донорских генов постепенно становятся разбавляется после каждого поколения роста бактерий до тех пор, пока донорский фенотип не сможет больше не выражаться. На селективной среде, на которой только бактерии с донорский фенотип может расти, абортивные трансдуктанты производят крошечные колонии, которые легко отличить от колоний стабильных трансдуктантов.Частота абортивной трансдукции обычно на один-два порядка больше чем частота генерализованной трансдукции, что указывает на то, что большинство клеток инфицированные генерализованными трансдуцирующими фагами, не продуцируют рекомбинантные потомство.
Специализированная трансдукция отличается от обобщенной трансдукции несколькими способами. Это опосредуется только определенными умеренными фагами и лишь несколькими конкретными донорами. гены могут передаваться бактериям-реципиентам. Специализированные трансдуцирующие фаги образуются только тогда, когда лизогенные донорские бактерии входят в литический цикл и высвобождают фаговое потомство.Специализированные фаги-трансдукторы — это редкие рекомбинанты, которые не имеют части нормального генома фага и содержат часть бактериального хромосома расположена рядом с местом прикрепления профага. Многие специализированные трансдуцирующие фаги являются дефектными и не могут завершить литический цикл фага рост в инфицированных клетках, если не присутствуют фаги-помощники, функции фага. Специализированная трансдукция является результатом лизогенизации бактерия-реципиент специализированным трансдуцирующим фагом и экспрессией донорские гены.Конверсия фагов и специализированная трансдукция имеют много сходства, но происхождение генов конверсии в умеренном климате фаги неизвестны.
Конъюгация
При конъюгации прямой контакт между донорскими и реципиентными бактериями приводит к установление цитоплазматического мостика между ними и перенос части или всего донорского генома реципиенту. Донорская способность определяется конкретными конъюгативные плазмиды называются плазмидами фертильности или половыми плазмидами.
Плазмида F (также называемая фактором F) E.coli — прототип для плазмид фертильности грамотрицательных бактерий. Штаммы E. coli с внехромосомной плазмидой F называются F + и функционируют как доноры, тогда как штаммы, лишенные Плазмиды F представляют собой F — и ведут себя как реципиенты. Спряжение функции плазмиды F определяются кластером из не менее 25 переносов ( tra ) гены, определяющие экспрессию F-пилей, синтез и перенос ДНК во время спаривания, вмешательство в способность F + бактерии для выполнения и других функций.Каждая бактерия F + имеет от 1 до 3 пилей F, которые связываются с определенным белок внешней мембраны (продукт гена ompA) на бактериях-реципиентах, чтобы инициировать вязка. Образуется межклеточный цитоплазматический мостик, и одна нить F плазмидная ДНК передается от донора к реципиенту, начиная с уникального источника и прогрессирует в направлении от 5 ‘до 3’. Переданный цепь превращается в кольцевую двухцепочечную плазмидную ДНК F у реципиента бактерии, и у донора синтезируется новая цепь для замены перенесенная прядь.Обе экзонъюгантные бактерии — F + , поэтому плазмида F может распространяться инфекцией среди генетически совместимых популяции бактерий. Помимо роли F-пилей в конъюгации, они также функционируют как рецепторы для донор-специфичных (специфичных для мужчин) фагов.
Плазмида F в E. coli может существовать как внехромосомная генетический элемент или быть интегрированным в бактериальную хромосому (). Поскольку плазмида F и Бактериальные хромосомы представляют собой обе кольцевые молекулы ДНК, взаимная рекомбинация между ними образуется более крупный круг ДНК, состоящий из вставленной плазмидной ДНК F линейно в хромосому. E. coli содержит несколько копий нескольких различных генетических элементов, называемых последовательностями вставки (см. транспозоны для более подробной информации), в различных местах в его хромосоме и в Плазмида F. Гомологичная рекомбинация между последовательностями вставки в хромосома и плазмида F приводит к преимущественной интеграции плазмиды F на хромосомных участках, где расположены последовательности вставки. Хромосомный сайты, где обнаруживаются последовательности вставки, различаются, однако, среди штаммов E.coli .
Рисунок 5-9
Роль плазмиды F в определении состояний донора и реципиента E. coli . Плазмида F представляет собой специфические конъюгативные плазмиды, которые контролировать донорскую способность E. coli . F- штаммы отсутствуют плазмиды F и являются генетическими реципиентами. F + штаммы гавани (подробнее …)
Штамм E. coli с интегрированной плазмидой F сохраняет свою способность выступать донором в супружеских отношениях. Поскольку донорский штамм с интегрированные факторы F могут передавать хромосомные гены реципиентам с высоким КПД, они называются Hfr ( H igh f Requency r ecombination) штаммов.Перенос одноцепочечной ДНК от донора Hfr к реципиенту начинается от источника внутри плазмиды F и происходит, как описано выше, за исключением того, что перенесенная ДНК является гибридной. репликон, состоящий из плазмиды F, интегрированной в бактериальную хромосому. Перенос всего этого репликона, включая бактериальную хромосому, требует примерно 100 минут. Идентичность первого хромосомного гена, который будет переносится и полярность хромосомного переноса определяется сайтом интеграции плазмиды F и ее ориентации относительно бактериальная хромосома.Потому что спаривающиеся бактерии обычно отделяются спонтанно до того, как вся хромосома будет перенесена, конъюгация обычно переносится только фрагмент донорской хромосомы в реципиент. Вероятность того, что донорский ген войдет в бактерию-реципиент во время уменьшения конъюгации, следовательно, как его расстояние от начала F (и, следовательно, время его передача) увеличивается. Спаривающиеся клетки также могут быть разделены экспериментально с помощью подвергая их сильным сдвигающим усилиям в механическом блендере; это называется прерванной вязкой.Формирование рекомбинантного потомства требует рекомбинация между перенесенной донорской ДНК и геномом реципиента бактерия. Анализ потомства от вязок, прерванных после разных интервалы демонстрируют, какие хромосомные гены передаются первыми конкретных донорских штаммов, последовательное время входа для генов, которые передаются впоследствии, и все более низкая вероятность того, что гены перенесенные позже появятся в рекомбинантном потомстве. Круглость генетическая карта E.coli был первоначально выведен из перекрывающиеся, циклически переставленные группы связанных генов, которые были перенесены рано отдельными донорскими штаммами, в которых фактор F был интегрирован на разные хромосомные места.
При скрещивании бактерий F + и F — , только плазмида F передается реципиентам с высокой эффективностью. Хромосомные гены передаются с очень низкой эффективностью, и это спонтанные мутанты Hfr в популяциях F + , которые опосредуют передача донорских хромосомных генов.В вязках между Hfr и F — штаммы , сегмент плазмиды F, содержащий tra область переносится последней, после всего бактериального хромосома перенесена. Большинство рекомбинантов от вязок между Hfr и F — клетки не могут наследовать весь набор плазмиды F. гены и фенотипически являются F —. В вязках между F + и F — штаммы , плазмида F быстро распространяется среди бактериальной популяции, и большинство рекомбинантов являются Факс + .
Интегрированные плазмиды F в штаммах Hfr иногда могут быть вырезаны из бактериального хромосома. Если вырезание полностью обращает процесс интегрирования, F + клетки производятся. Однако в редких случаях иссечение происходит путем рекомбинации с участием инсерционных последовательностей или других генов на бактериальном хромосомы, расположенные на некотором расстоянии от исходного сайта интеграции. В таких случаях сегменты бактериальной хромосомы могут включаться в гибридные плазмиды F, которые называют плазмидами F ‘(см.).Подобными процессами сегменты бактериального хромосома иногда может быть включена в плазмиды R с образованием гибридных R ‘плазмиды. Конъюгативные плазмиды R ‘могут функционировать как плазмиды фертильности, потому что они могут интегрироваться в бактериальную хромосому путем гомологичной рекомбинации и опосредуют передачу хромосомных генов во время спаривания с бактериями-реципиентами. F ‘ плазмиды, R ‘плазмиды, специализированные трансдуцирующие фаги и рекомбинантные плазмиды или фаги, сконструированные путем клонирования генов (описанные ниже), представляют собой гибридные репликоны которые могут включать сегменты бактериальной хромосомы.Следовательно, любой из этих генетические элементы могут быть использованы для создания частично диплоидных бактериальных штаммы, необходимые для тестов на комплементарность и других целей.
Конъюгация также происходит у грамположительных бактерий. Грамположительные донорские бактерии вырабатывают адгезины, которые заставляют их агрегироваться с реципиентными клетками, но половые пили не участвуют. У некоторых видов Streptococcus , реципиент бактерии производят внеклеточные половые феромоны, которые вызывают изменение фенотипа донора. экспрессироваться бактериями, несущими соответствующую конъюгативную плазмиду, и конъюгативная плазмида не дает донорским клеткам продуцировать соответствующие феромон.
Рекомбинация
Рекомбинация включает разрыв и соединение родительских молекул ДНК с образованием гибридные, рекомбинантные молекулы. Было обнаружено несколько различных видов рекомбинации. идентифицированы, которые зависят от различных характеристик участвующих геномов и требуют активности различных генных продуктов. Специфические ферменты, которые действуют на ДНК (например, экзонуклеазы, эндонуклеазы, полимеразы, лигазы) участвуют в рекомбинации. Подробное обсуждение биохимических событий в рекомбинации выходит за рамки данной главы.
Обобщенная рекомбинация включает молекулы донорной и реципиентной ДНК, которые имеют гомологичные нуклеотидные последовательности. Взаимные обмены могут происходить между любыми гомологичные сайты-доноры и реципиенты. В E. coli продукт гена recA важен для генерализованной рекомбинации, но и другие генные продукты тоже участвуют.
Сайт-специфическая рекомбинация включает взаимные обмены только между определенными сайты в молекулах донорской и реципиентной ДНК. Ген recA продукт не требуется для сайт-специфической рекомбинации.Интеграция умеренный бактериофаг l в хромосому E. coli хорошо изученный пример сайт-специфической рекомбинации (). Конкретные сайты вложений (att) на Хромосома E. coli и ДНК фага λ имеют общие основная последовательность из 15 нуклеотидов, внутри которой происходит реципрокная рекомбинация, фланкированы соседними последовательностями, не гомологичными фаговым и бактериальным геномы. В фаге λ продукт гена int (интеграза) требуется для специфичного для сайта интеграционного события в лизогенизация; продукты int и Гены xis (эксцизионаза) необходимы для дополнительное сайт-специфическое иссечение, которое происходит во время индукции литического развитие фага в лизогенных клетках.
Рисунок 5-10
Интеграция и удаление бактериофага λ примеры сайт-специфической рекомбинации. λ ДНК показана тонкими линиями, а хромосомная ДНК — толстыми линиями. линий. Места прикрепления (атт) представляют собой закрытые ящики для бактериального хромосома и открытые боксы (подробнее …)
Незаконная рекомбинация — это термин, используемый для описания негомологичных, аберрантных рекомбинационные события, такие как те, которые участвуют в формировании специализированных трансдуцирующие фаги. Механизмы нелегитимной рекомбинации: неизвестный.
Транспозоны
Транспозоны — это сегменты ДНК, которые могут перемещаться из одного участка молекулы ДНК в другие сайты-мишени в той же или другой молекуле ДНК. Процесс называется транспозицией и происходит с помощью механизма, который не зависит от обобщенная рекомбинация. Транспозоны являются важными генетическими элементами, потому что они вызывают мутации, опосредуют геномные перестройки, функционируют как переносимые области генетической гомологии, приобретают новые гены и вносят свой вклад в их распространение среди бактериальных популяций.Часто введение транспозона прерывает линейную последовательность гена и инактивирует его. Транспозоны имеют важную роль в возникновении делеций, дупликаций и инверсий сегментов ДНК, как а также слияния репликонов. Транспозоны не являются самовоспроизводящимися генетическими элементы, однако, и они должны интегрироваться в другие репликоны для поддержания стабильно в бактериальных геномах.
Большинство транспозонов имеют ряд общих черт. Каждый транспозон кодирует функции, необходимые для его транспозиции, в том числе фермент транспозазы, который взаимодействует со специфическими последовательностями на концах транспозона.В течение при транспозиции короткая последовательность целевой ДНК дублируется, и транспозон вставлен между непосредственно повторяющимися целевыми последовательностями. Длина этого короткая дупликация варьирует, но характерна для каждого транспозона. В Предполагается, что дупликация включает асимметричное расщепление ДНК в мишени. сайт с последующим синтезом новых комплементарных цепей, соответствующих область между сайтами расщепления. Некоторые транспозоны вставляются практически в любые целевая последовательность, тогда как другие имеют относительно строгую целевую специфичность.Различают два типа транспозиции. Удаление транспозона из донорский сайт с последующей его вставкой в целевой сайт называется нерепликативным. транспозиция. Если транспозон на донорском сайте реплицируется и копия вставлен в целевой сайт, однако процесс называется репликативным транспозиция. Процесс репликативной транспозиции может включать образование коинтеграция, одиночная кольцевая молекула ДНК, состоящая из двух репликонов, соединенных с копиями транспозона в чередующейся последовательности.Резолюция коинтеграция в составляющие его репликоны часто достигается за счет кодируемая транспозоном резольваза, которая катализирует сайт-специфическую рекомбинацию между транспозоны. Обобщенная рекомбинация между гомологичными транспозонами может также приводят к образованию или расслоению коинтегратов. Транспозиция отличается от сайт-специфической рекомбинации путем дублирования сегмента целевой последовательности и с использованием множества различных целевых последовательностей для одного донора последовательность.
Большинство транспозонов в бактериях можно разделить на три основных класса ().Последовательности вставки и связанные составные транспозоны составляют первый класс. Последовательности вставки самые простые в структуре и кодируют только функции, необходимые для транспонирования. Известный Последовательности вставки варьируются по длине примерно от 780 до 1500 нуклеотидов. пары, имеют короткие (15-25 пар оснований) перевернутые повторы на концах и не являются тесно связаны друг с другом. ДНК между инвертированными концевыми повторами содержит один (или редко два) гена транспозазы и не кодирует резольвазу.Сложные транспозоны различаются по длине от примерно 2000 до более чем 40000. пары нуклеотидов и содержат последовательности вставки (или близкородственные последовательности) на каждом конце, обычно в виде перевернутых повторов. Весь сложный элемент может транспонировать как единое целое. ДНК между концевыми вставками последовательностей комплекса транспозоны кодируют несколько функций, которые не важны для транспозиции. У важных с медицинской точки зрения бактерий гены, определяющие приверженность к лечению. антигенов, токсинов или других факторов вирулентности, или указать устойчивость к одному или больше антибиотиков, часто находятся в сложных транспозонах.Известные примеры сложных транспозонов Tn 5 и Тн 10 , определяющие устойчивость к канамицину и тетрациклин соответственно. Сложные транспозоны, вероятно, эволюционируют транспозиция гомологичных последовательностей вставок в близлежащие сайты в ДНК молекула.
Рисунок 5-11
Характеристики репрезентативных транспозонов (жирные линии) интегрированы в бактериальную хромосому (тонкие морщинки). Транспозоны являются важными генетическими элементами, потому что они вызывают мутации, опосредуют геномные перестройки, функционируют как переносимые области генетического (подробнее…)
Второй класс транспозонов состоит из высокогомологичных ТН Семейство . Эти транспозоны имеют более длинные (от 35 до 40 пара оснований) концевые инвертированные повторы, чем описанные сложные транспозоны выше, но в них отсутствуют концевые инсерционные последовательности. Все члены семьи кодируют функции транспозазы и резольвазы. Хорошо известные примеры из Tn Семейство транспозонов включает ампициллин резистентность транспозона Tn 3 и Tn 1000 (гамма-дельта транспозон), обнаруженный в F плазмида.Семья TnA занимает важное место в истории медицины. микробиология. Развитие высокой устойчивости к ампициллину у Haemophilus influenzae и Neisseria gonorrhoeae в 1970-е годы, что сильно ограничивало полезность ампициллина для лечения гонореи и гемофильной инфекции в регионах там, где такие штаммы стали преобладать, было вызвано распространением ампициллина детерминанты устойчивости из транспозонов Tn A в плазмидах Enterobacteriaceae к плазмидам в Haemophilus и Нейссерия .
Третий класс транспозонов состоит из бактериофага Mu и родственных ему умеренных фаги. Весь геном фага функционирует как транспозон, и репликация ДНК фага во время вегетативного роста происходит путем репликативной транспозиции. Интеграция профагов может происходить во многих разных участках бактериального хромосома и часто вызывает мутации. По этой причине Mu и родственные фаги иногда их называют фагами-мутаторами.
Четвертый класс транспозонов, обнаруженный у грамположительных бактерий и представлен Тн 917 , состоит из конъюгативных транспозонов которые полностью отличаются от транспозонов, описанных выше.В конъюгативный транспозон не создает дупликации целевой последовательности в которую он вставляется, а у грамположительных бактерий штамм-хозяин, несущий транспозон может действовать как супружеский донор. Бактерии-реципиенты не нужно близкородственен донорской бактерии. Транспозон иссекают из хромосома донора и передается путем конъюгации реципиенту, где он случайным образом интегрируется в хромосому. Tn 917 кодирует устойчивость к тетрациклину, но другие более крупные конъюгативные транспозоны могут кодировать дополнительная устойчивость к антибиотикам.Конъюгативные транспозоны, по-видимому, являются основным причина распространения устойчивости к антибиотикам у грамположительных бактерий.
Некоторые роли транспозонов в эволюции бактерий проиллюстрированы рассмотрением кишечные грамотрицательные бактерии и строение их плазмид. Бактерии собранные в доантибиотическую эру, содержали много плазмид, но они обычно отсутствовали детерминанты устойчивости. Многие из плазмид R из текущего клинические изоляты принадлежат к тем же группам несовместимости, что и обнаруженные плазмиды ранее, но они также определяют устойчивость к нескольким антибиотикам.В близкие отношения между их репликонами убедительно доказывают, что многие текущие плазмиды R произошли от более старых плазмид путем приобретения устойчивости детерминанты. Некоторые из плазмид с множественной устойчивостью к антибиотикам имеют индивидуальные транспозоны с несколькими детерминантами устойчивости, другие имеют несколько транспозоны резистентности расположены на разных участках, а третьи содержат сложные гибридные транспозоны устойчивости, образованные путем интеграции одного транспозона в другой. Поэтапное получение детерминант сопротивления может привести к в некоторых случаях к образованию составных транспозонов, кодирующих несколько детерминанты устойчивости.Терапевтическое использование антибиотиков и их включение в корм для животных обеспечивает селективные преимущества для бактерий с плазмидами R, тогда как конъюгация, трансформация и трансфекция обеспечивают средства для распространение плазмид R внутри и между видами бактерий. После плазмида, несущая транспозон, вводится в нового бактериального хозяина, транспозон и его детерминанты могут прыгать в хромосому или коренные плазмиды нового хозяина. Следовательно, стабильность мобилизующей плазмиды в новый бактериальный хозяин не важен для сохранения генетических детерминант расположен на транспозоне.
Рекомбинационная ДНК и клонирование генов
Существует множество методов создания гибридных молекул ДНК in vitro (рекомбинантная ДНК) и охарактеризовать их. Такие методы включают выделение определенных генов в гибридных репликоны, определяя их нуклеотидные последовательности и создавая мутации в обозначенные места (сайт-направленный мутагенез). Клон — это популяция организмы или молекулы, полученные в результате бесполого размножения от одного предка. Ген клонирование — это процесс включения чужеродных генов в репликоны гибридной ДНК.Клонированные гены могут быть экспрессированы в соответствующих клетках-хозяевах, и фенотипы, которые они определяют, что могут быть проанализированы. Некоторые ключевые концепции, лежащие в основе репрезентативных методов резюмируются здесь.
Первым шагом в клонировании генов является получение фрагментов донорской ДНК механическим или ферментативные методы. Некоторые эндонуклеазы рестрикции, обозначенные как класс II, являются особенно полезно для получения определенных фрагментов молекул ДНК. Они рассекаются обе нити двухцепочечных молекул ДНК в определенных палиндромных последовательностях (сайты рестрикции), длина которых обычно варьируется от четырех до восьми нуклеотидов, и полученные фрагменты ДНК называются рестрикционными фрагментами.Некоторое ограничение эндонуклеазы расщепляют совпадающие сайты с образованием фрагментов ДНК с тупыми концами, и другие разрезают в шахматном порядке для создания фрагментов ДНК с короткими, самокомплементарные одноцепочечные 5 ‘или 3’ концы (см.). Случайная вероятность того, что n соседние нуклеотиды в цепи ДНК будут соответствовать определенному сайту рестрикции составляет примерно 1/4 n . Сайты для ферментов, распознающих уникальные 4, 6 или 8 нуклеотидные мишени могут встречаться примерно один раз из каждых 256, 4096 или 65 536 нуклеотиды соответственно.Выбирая подходящие рестрикционные ферменты, специфическая ДНК молекулы, включая бактериальные хромосомы, плазмиды и геномы фагов, могут быть переваривается в наборы ограничивающих фрагментов, которые имеют соответствующие размеры для конкретных Приложения.
Таблица 5-3
Особенности ограничения типичного класса II Эндонуклеазы.
Карта ограничений определяет позиции целевых сайтов для конкретного ограничения эндонуклеазы в молекуле ДНК. Карты ограничений доступны для многих клонированных ДНК. фрагментов, плазмид и геномов фагов, а также для всей хромосомы E.coli и несколько других бактерий.
Второй этап клонирования генов — создание гибридных репликонов, состоящих из донорской ДНК. фрагменты и вектор клонирования (). Клонирующие векторы представляют собой небольшие репликоны плазмид или фагов, которые имеют один или несколько сайты рестрикции, в которые может быть вставлена чужеродная ДНК. Гибридные репликоны производится с использованием ДНК-лигазы для соединения рестриктированной векторной ДНК с донорной ДНК фрагменты, которые имеют совместимые концы, или, альтернативно, синтетические олигонуклеотиды используются в качестве линкеров для создания совместимости между донорскими и векторными молекулами ДНК с разными концами.Лигирование вектора с гетерогенным набором фрагментов ДНК из донорский геном называется клонированием дробовика, а сбор рекомбинантной ДНК молекулы, содержащие различные фрагменты, называются геномной библиотекой. Если специфический фрагмент ДНК доступен, он может быть включен в рекомбинантный репликон путем прямого клонирования в соответствующий вектор, выбранный из широкого разнообразия доступных векторов. Плазмидные и фаговые векторы используются в основном для клонирования небольших вставляет обычно меньше 10 кб. Примеры других векторов специального назначения включают космиды, которые представляют собой плазмидные векторы, которые могут быть упакованы в фаговые капсиды (лямбда космиды принимают вставки размером до 30-40 т.п.н.) и фагмиды, которые являются плазмид-фагами. гибридные репликоны, которые могут существовать в виде плазмид или одноцепочечных ДНК-фагов в разных условиях эксперимента.Космиды фага P1 могут принимать вставки до 100 т.п.н., а молекулы ДНК еще большего размера можно клонировать в дрожжевые искусственные хромосомы (YAC), которые могут стабильно поддерживать вставки размером до и более 1 Мбит / с в размер. Другие специализированные векторы обнаруживают промоторы, сигналы терминации транскрипции, или другие регуляторные элементы во вставках чужеродной ДНК или, наоборот, обеспечивают промоторы, с которых может быть инициирована транскрипция клонированных генов.
Рисунок 5-12
Схематическое изображение эксперимента по клонированию гена.Плазмидный вектор клонирования pBR322 имеет размер 4,36 пар оснований, имеет гены на устойчивость к ампициллину (amp r ) и тетрациклину ( tet r ) и имеет только один HindIII сайт рестрикции, расположенный внутри (подробнее …)
Заключительными этапами клонирования генов является введение гибридных репликонов в соответствующие клетки-реципиенты и проверить их на экспрессию интересующих донорских генов. Прокариотический клетки (включая бактерии) или эукариотические клетки (включая дрожжи, животные или растения) ячеек) могут использоваться в качестве получателей, но они различаются по своему вседозволенность для определенных репликонов, транскрипционные сигналы, которые они распознают и посттрансляционные модификации белковой структуры, которые они может выполнить.Можно вводить рекомбинантные молекулы ДНК, полученные in vitro. непосредственно в клетки-реципиенты путем трансформации или трансфекции. Кроме того, клоны в космидных или фаговых векторах могут быть упакованы в фаговые оболочки и введены в восприимчивые клетки-реципиенты путем трансдукции. Используя специализированные векторы (челнок векторов), которые могут реплицироваться в нескольких типах клеток, гены любого организма могут быть клонированы и обработаны в удобной бактериальной системе, а затем повторно введены в клетки исходного организма для анализа в их естественных окружающая обстановка.
Существует множество методов идентификации бактерий, содержащих рекомбинантную ДНК. молекулы. Большинство векторов клонирования имеют гены признаков, которые могут быть положительно выбран, например, устойчивость к антибиотикам. Кроме того, часто можно ввести чужеродную ДНК в вектор клонирования в сайт, который инактивирует несущественная, но легко узнаваемая векторная функция. Если оба эти условия выполняются, бактерии, содержащие рекомбинантные молекулы, могут быть отобраны и легко отличить от бактерий, содержащих только вектор.Бактерии в геномная библиотека, содержащая конкретный клонированный ген, может быть идентифицирована с помощью биохимические или иммунологические методы тестирования желаемого генного продукта. В качестве альтернативы интересующий клонированный ген может быть обнаружен непосредственно с помощью нуклеиновой кислоты. методы кислотной гибридизации при условии, что доступен конкретный ДНК- или РНК-зонд. Поскольку вставка чужеродной ДНК в вектор клонирования в соответствующем сайте делает не инактивировать его способность реплицироваться в соответствующих клетках-реципиентах, гибрид представляющие интерес репликоны могут быть амплифицированы путем репликации, и рекомбинантная ДНК молекулы или их генные продукты могут быть очищены и изучены.Способность очищать специфические молекулы ДНК сделали возможным развитие ферментативных и химических методов для определения их нуклеотидных последовательностей и современные методы введения мутации в определенных сайтах в клонированных генах основаны на знании их ограничения карты или нуклеотидные последовательности.
Методы рекомбинантной ДНК позволяют клонировать определенные фрагменты ДНК из любых источника в векторы, которые могут быть изучены на хорошо охарактеризованных бактериях, в эукариотические клетки, или in vitro .Применения клонирования ДНК быстро расширяется во всех областях биологии и медицины. В медицинской генетике такие область применения варьируется от пренатальной диагностики наследственных заболеваний человека до характеристика онкогенов и их роль в канцерогенезе. Фармацевтическая приложения включают крупномасштабное производство из клонированных человеческих генов биологических продукты с терапевтической ценностью, такие как полипептидные гормоны, интерлейкины и ферменты. Приложения в общественном здравоохранении и лабораторной медицине включают разработки вакцин для предотвращения конкретных инфекций и зондов для диагностики конкретных инфекции путем гибридизации нуклеиновых кислот или полимеразной цепной реакции (ПЦР).В в последнем процессе используются олигонуклеотидные праймеры и ДНК-полимераза для амплификации специфических последовательности ДНК-мишени во время нескольких циклов синтеза in vitro, что делает его возможно обнаружение редких целевых последовательностей ДНК в клинических образцах с большим чувствительность.
Регуляция экспрессии генов
Фенотипические свойства бактерий определяются их генотипами и ростом условия. Для бактерий в чистой культуре часто возникают изменения условий роста. в предсказуемых физиологических адаптациях у всех членов популяции.Как правило, основные генные продукты производятся в количествах, обеспечивающих самый быстрый рост данной среде, и продукты, необходимые при особых обстоятельствах, производятся только тогда, когда они нужны.
Физиологические адаптации часто связаны с изменениями метаболической активности. Поток метаболитов через определенные биохимические пути можно контролировать. как путем регулирования синтеза определенных ферментов, так и путем изменения активности существующих ферментов. Механизмы, регулирующие экспрессию генов, воздействуя на здесь обсуждается синтез конкретных генных продуктов.
Специфическая регуляция включает ген или группу генов, участвующих в конкретном метаболический процесс. Индукция и репрессия позволяют бактериям регулировать производство конкретных генных продуктов в ответ на соответствующие сигналы. Обычно катаболический ферменты индуцируются, когда субстрат для пути роста присутствует в ростовой среда, и биосинтетические ферменты подавляются продуктом пути. Ферменты, которые участвуют в одном биохимическом пути, часто занимают соседние положения на бактериальной хромосоме и координированно индуцируются или подавляются.Они образуют оперон, группу смежных генов, которые транскрибируются как единое целое. единица и переводится для получения соответствующих генных продуктов. Организация в оперон — важная стратегия для скоординированной регуляции экспрессии генов в бактериях. Опероны, которые могут быть индуцированы или подавлены, контролируются связыванием специфические регуляторные белки к определенным нуклеотидным последовательностям, которые функционируют как регуляторные сайты внутри оперона. Сравнение аминокислотных последовательностей многих этих различных регуляторных белков показали, что они могут быть сгруппированы вместе в семейства регуляторов (например,г. семейство белков lysR), которые могли развиться от общих предковых генов. Члены семейства lysR включают регуляторы таких разнообразные явления, такие как метаболизм лизина, цистеина и метионина у E. coli и репрессия железа в V. cholerae .
Глобальная регуляция одновременно изменяет экспрессию группы генов и оперонов, вместе называемые регулонами, которые контролируются одним и тем же регуляторным сигналом. Глобальное регулирование определяет реакцию бактерий на основные питательные вещества, такие как углерод, азот или фосфат, реакции на стрессы, такие как повреждение ДНК или тепло шока, а также синтез патогенами специфических факторов вирулентности при росте в их животные-хозяева.Количество определенного белка в бактериальной клетке может варьироваться. от нуля до многих тысяч молекул. Этот широкий диапазон часто определяется комбинированное действие нескольких регуляторных механизмов, влияющих на экспрессию соответствующий структурный ген. Регулирование достигается путем определения того, как часто ген транскрибируется в функциональную мРНК, насколько эффективно транслируется мРНК в белок, как быстро разлагается мРНК, как быстро белковый продукт переворачивается, и может ли активность белкового продукта быть изменена аллостерические эффекты или ковалентные модификации.
мРНК как транскрипционные единицы
Экспрессия гена начинается с ДНК-зависимой РНК-полимеразы (РНК-полимеразы) катализирует транскрипцию специфической мРНК из одной цепи ДНК-матрицы. Связывание РНК-полимеразы с ДНК происходит в определенных сайтах, называемых промоторами, и транскрипция начинается рядом с промотором. Сильные промоутеры могут взаимодействовать эффективно с РНК-полимеразой и инициировать транскрипцию с высокой скоростью; слабый промоторы инициируют транскрипцию с медленной скоростью. В любом случае мРНК синтезируется от его 5′-конца к его 3′-концу на приблизительно с постоянной скоростью, пока РНК-полимераза не распознает другой специфический сайт назвал терминатором.Затем РНК-полимераза отделяется от матрицы и транскрипция мРНК завершена.
Отдельные молекулы мРНК могут кодировать один или несколько полипептидов. Транскрипция оперон продуцирует полицистронную мРНК, которая кодирует несколько полипептидов. Трансляция полицистронных мРНК приводит к координированному синтезу кодируемых полипептиды, но каждый полипептид синтезируется как отдельная молекула. А специфический сайт связывания рибосомы расположен прямо перед началом каждого кодирующая последовательность на молекуле мРНК.
Информационные РНК в бактериях быстро разлагаются со средним периодом полураспада несколько минут, в отличие от тРНК и рРНК, которые намного более стабильны. Хотя мРНК составляют около половины вновь синтезированной РНК, они представляют собой только небольшая часть общей РНК. Короткий период полужизни мРНК имеет важные последствия для экспрессии генов. Если синтез определенной мРНК предотвращается, продукция соответствующих полипептидов быстро снижается.
Контроль экспрессии генов осуществляется путем регулирования одного или нескольких этапов путь от матрицы ДНК к продукту активного гена.Одновременный регулирование на нескольких уровнях позволяет лучше контролировать экспрессию генов, чем было бы возможно с помощью единого регулирующего механизма. Самый распространенный способ регулировать экспрессию генов в бактериях — контролировать производство специфических мРНК. Поскольку скорость удлинения молекулы РНК приблизительно равна константа, основными факторами, контролирующими синтез мРНК, являются скорость инициации и вероятность того, что полный текст будет произведено.
Регуляция инициации транскрипции
Некоторые мРНК в бактериях синтезируются с постоянной скоростью, что приводит к конститутивная продукция кодируемых полипептидов.Количество конкретных мРНК и полипептиды, продуцируемые разными конститутивными генами, сильно различаются, однако и часто отражают различия в силе сторонников тех гены.
Транскрипция многих оперонов регулируется в ответ на изменение окружающей среды. условия. Промоторы определяют максимальную скорость инициации транскрипции. для таких оперонов, но регуляторные белки участвуют в контроле транскрипция. Нуклеотидные последовательности в оперонах, к которым относятся специфические регуляторные связывание белков называется регуляторными сайтами или операторами.Операторы и промоутеры расположены близко друг к другу внутри оперонов и могут иметь перекрывающиеся ДНК последовательности. Связывание регуляторных белков с операторами может увеличиваться (положительное регулирование) или уменьшение (отрицательное регулирование) частоты инициация транскрипции. Белки, которые функционируют как негативные регуляторы: обычно называемые репрессорами. Поскольку регуляторные белки могут диффундировать через цитоплазма, структурные гены регуляторных белков не обязательно должны быть связаны к целевым оперонам.
Способность определять присутствие или отсутствие определенных соединений и изменять темпы синтеза соответствующих генных продуктов имеют центральное значение для контроля экспрессия гена. Регуляторные белки предлагают одно решение этой проблемы связь стимул-реакция. Многие регуляторные белки бифункциональны и связываются не только к соответствующим операторам, но и к конкретным эффекторам, которые небольшие молекулы, такие как определенные сахара, аминокислоты и другие метаболиты. Кроме того, регуляторные белки являются аллостерическими, что означает, что они могут существовать в различные конформации, которые проявляют различную аффинность связывания для своих родственные операторы и эффекторы.Достаточная концентрация эффекторных благовоний образование регуляторного комплекса белок-эффектор, который имеет либо высокий, либо низкое сродство к оператору в каждом конкретном случае. В отрицательно регулируемом систем, эффектор функционирует как корепрессор, если регулирующий белок-эффекторный комплекс является активным репрессором, а эффекторный комплекс функционирует как индуктором и вызывает дерепрессию, если свободный регуляторный белок является активным репрессор. Напротив, в положительно регулируемых системах эффектор стимулирует экспрессия оперона, если регуляторный белок-эффекторный комплекс представляет собой положительный регулятор, а эффектор подавляет экспрессию оперона, если свободный регуляторный белок является положительным регулятором.
Лактозный (lac) оперон E. coli является примером индуцибельный, отрицательно регулируемый оперон (). Ген lacI кодирует репрессор, который связывается с оператором lac и предотвращает транскрипцию из Промотор lac . Структурный ген этого репрессора отдельно от lac-оперона, и репрессор синтезируется конститутивно при низкая ставка. Когда индуктор связывается с lac-репрессором, комплекс не может связываться с оператора и не может предотвратить связывание РНК-полимеразы с промотором.При других благоприятных условиях экспрессируется оперон lac , в результате синтеза β-галактозидазы, β-галактозида пермеаза и β-галактозид трансацетилаза. Модель lac оперон может быть индуцирован лактозой или структурно родственными соединениями, такими как изопропил-b-D-тиогалактозид (IPTG). IPTG называют безвозмездным индуктором потому что он индуцирует lac-оперон, но не является субстратом для β-галактозидаза. Отрицательная регуляция также встречается во многих биосинтетических оперонов в E.coli . В таких оперонах продукт биосинтетический путь действует как эффектор для негативных регуляторных система.
Рисунок 5-13
Регулирование оперона lac в E. coli . Структурные гены lacZ , lacY и код lacA для β-галактозидазы, β-галактозид пермеаза и β-галактозид трансацетилаза, соответственно. Физиологическая роль lacA заключается в неизвестный. Репрессор lac (подробнее…)
Оперон арабинозы ( ara ) в E. coli является как положительно и отрицательно регулируется. В присутствии арабинозы нормативный белок стимулирует транскрипцию оперона ara. В отсутствие арабинозы однако регуляторный белок репрессирует оперон ara .
Опероны часто управляются более чем одним механизмом. Когда E. coli выращивают в среде, содержащей глюкозу и альтернативный источник углерода, такой как лактоза или арабиноза, индукция lac или ara оперон и использование лактоза или арабиноза задерживаются до тех пор, пока не будет израсходована глюкоза.Этот явление называется диуксическим ростом. Неспособность вызвать оперон lac или ara в присутствии глюкозы является примером репрессии катаболита. Лак и ara опероны положительно регулируются циклический-3 ‘, 5’-аденозинмонофосфат (цАМФ) и белок активатора гена катаболита (CAP) (продукт гена crp). В Комплекс цАМФ-ЦАП взаимодействует с сайтами связывания ЦАП в регуляторных областях некоторые опероны, в том числе опероны lac и ara, и стимулирует транскрипцию от соответствующих промоутеров.Уровень внутриклеточного цАМФ у E. coli высокий во время роста в отсутствие глюкозы и низкий во время роста в присутствии глюкозы. Катаболиты репрессии должны, следовательно, к отсутствию активации цАМФ-зависимых оперонов, когда бактерии выращены в присутствии глюкозы или другого быстро метаболизируемого углерода источники.
Регуляция терминации транскрипции
Аттенуация — это механизм регуляции оперонов путем прекращения транскрипции мРНК преждевременно.Аттенуация часто встречается в биосинтетических оперонах, включая trp , гистидин ( his ), треонин ( th ), изолейцин-валин ( ilv ) и фенилаланиновые (phe) опероны. Оперон trp в E. coli контролируется как репрессией, так и ослаблением. в наличие избытка триптофана, инициация транскрипции с trp Промотор репрессирован. Кроме того, однако, те транскрипты, инициированные с промотора trp , являются обычно заканчивается раньше любого из структурных генов Расшифровано trp, оперонов.Концентрация внутриклеточный триптофан, необходимый для поддержания репрессии, превышает уровень, необходимый для затухание. Такой двойной контроль позволяет клетке точно настроить выражение оперон trp в ответ на снижение концентрации триптофан.
Вторичная структура мРНК играет важную роль в механизме затухание. Все мРНК имеют лидерную последовательность между началом транскрипции. сайт и начало кодирующей последовательности для первого структурного гена.Для аминокислотные биосинтетические опероны, которые подвергаются аттенуации, лидер мРНК Последовательность имеет две отличительные особенности. Он кодирует короткий пептид, содержащий аминокислота, продуцируемая регулируемым путем, и она может образовывать альтернативу, взаимно несовместимые, двухцепочечные структуры РНК, которые участвуют в нормативные события. Например, пептид, кодируемый trp Лидерная последовательность мРНК содержит два соседних остатка триптофана, а пептид кодируемая лидерной последовательностью мРНК , имеет серию из семи последовательные остатки гистидина.показывает оперон trp и иллюстрирует альтернативный вторичный структур в лидерной последовательности trp мРНК . Есть три возможные вторичные структуры для этого региона, называемые сайтом паузы (сегменты 1 + 2), антитерминатор (сегменты 2 + 3) и аттенюатор (сегменты 3 + 4). Сегмент 1 сайта паузы перекрывается кодировкой область для пептида trpL . Какие второстепенные конструкции формируется зависит от эффективности трансляции trpL пептид.Когда сегменты 1 и 2 расшифровываются, они немедленно отжигаются и вызвать временную остановку РНК-полимеразы. Последующее инициирование трансляция пептида trpL нарушает сайт паузы и позволяет РНК-полимеразе продолжать транскрипцию. Если присутствует триптофан, транскрипция сегментов 3 и 4 и формирование структуры аттенюатора происходит, когда рибосома блокирует сегмент 2, в результате чего РНК-полимераза прекратить транскрипцию. Однако при дефиците триптофана триптофанил-тРНК также недостаточен, и рибосома останавливается на кодонах триптофана в сегменте 1.Это позволяет сегменту 2 отжигаться с вновь синтезированным сегментом 3, чтобы сформировать антитерминатор, тем самым делая сегмент 3 недоступным для отжига с сегментом 4. Таким образом предотвращается образование аттенюатора, и РНК-полимераза транскрибирует весь оперон trp . Таким образом истощение триптофан (фактически поставка триптофанил-тРНК) связан с регуляцией транскрипции биосинтетического оперона триптофана.
Рисунок 5-14
Регулировка оперона trp в E.coli . Организация оперона trp показана на верх фигуры. Пять структурных генов trpE , trpD , trpC , trpB и trpA кодируют ферменты которые катализируют терминальную последовательность реакций в триптофане формирование. (подробнее …)
В E. coli транскрипция и трансляция функционально спаренный. Бессмысленные мутации, вызывающие преждевременное прекращение трансляции часто вызывают снижение транскрипции более дистальных генов в том же опероне.Это явление называется полярностью. Рибосомы обычно инициируют трансляцию рост молекулы мРНК до завершения транскрипции, и такая трансляция маскирует сайты, которые в противном случае привели бы к прекращению действия РНК-полимеразы транскрипция. Преждевременное прекращение трансляции бессмысленным кодоном отделяет рибосомы от мРНК и позволяет РНК-полимеразе взаимодействовать с немаскированными сайтами терминации транскрипции.
В некоторых биологических системах, включая фаг лямбда, антитерминация используется как положительный регуляторный механизм для контроля экспрессии генов.Незамедлительно после заражение E. coli лямбда, РНК-полимераза связывается с двумя промоторов в лямбда-ДНК и инициирует дивергентные первичные транскрипты, которые оканчиваются на определенных сайтах лямбда-генома. Белок, кодируемый одним из первичные транскрипты взаимодействуют с РНК-полимеразой и позволяют ей продолжать транскрипция через первичные сайты терминации, тем самым выражая второй набор лямбда-генов. Один из продуктов, закодированный вторичной расшифровкой блокирует терминацию другой мРНК и активирует экспрессию третьего набора гены.Следовательно, антитерминация играет ключевую роль в управлении каскадом экспрессия гена во время литического роста фага лямбда. Антитерминация также участвует в регуляции оперонов рРНК E. coli.
Регуляция трансляции
Сайт связывания рибосомы на мРНК комплементарен последовательности на 3′-конец 16S рРНК. Взаимодействие между этими последовательностями облегчает формирование комплекса инициации синтеза белка. Как степень гомология с 16S рРНК и расстояние сайта связывания рибосомы от кодон инициации влияет на эффективность инициации трансляции.Использование кодонов в мРНК также влияет на эффективность трансляции. Информационные РНК для белков, которые требуются в больших количествах, как правило, используют кодоны, которые транслируются наиболее многочисленные виды тРНК, и обратное тоже верно.
Трансляционный контроль важен для регуляции синтеза рибосомных белки. Производство рибосом связано с высокими метаболическими затратами для бактерий, и при высоких скоростях роста рибосомы могут составлять почти половину клетки. масса. Большинство рибосомных белков и рРНК собраны в рибосомы, и пул свободных субъединиц рибосом очень мал.Гены рибосом белки организованы в несколько оперонов. Некоторые из свободных рибосом белки напрямую ингибируют трансляцию полицистронных мРНК, которые кодируют их, тем самым обеспечивая баланс синтеза рибосомальных белков с требование к их утилизации.
Регулоны и белки, передающие сигнал
Реглон — это группа генов или оперонов, контролируемых общим регулятором. Там есть несколько преимуществ в том, чтобы поставить под контроль разные опероны одним и тем же регулятор.Это позволяет связать восприятие одиночного стимула с экспрессия большого количества генов, которые могут быть необходимы для соответствующего ответ, и это устраняет потребность в скоординированно регулируемых генах быть сцепленным на бактериальной хромосоме. Стимул, к которому регулон Ответы могут быть внутриклеточным компонентом или сигналом окружающей среды. Отдельные опероны также могут регулироваться несколькими различными механизмы и выражаются в условиях, отличных от тех, которые влияют на весь регулон.
Более 40 различных регулонов были идентифицированы в E. coli . Конкретные примеры регулонов, которые отвечают на внутриклеточные компоненты, включают: описанный ранее регулон цАМФ-CAP и регулоны, контролируемые строгий ответ и ответ SOS. Когда рибосомы сталкиваются с незаряженной тРНК молекул во время синтеза белка активируется строгий ответ и приводит к быстрому прекращению синтеза рРНК. Новый нуклеотид под названием гуанозин-3′-дифосфат-5′-дифосфат (ppGpp) накапливается во время аминокислотного голодания.PpGpp, продуцируемый холостыми рибосомами, кажется, быть посредником строгого ответа, но точный механизм, вызывающий ингибирование синтеза рРНК неизвестно. Ответ SOS связан с повреждение ДНК и включает индукцию более 20 генов, участвующих в нескольких Пути репарации ДНК. Продукт гена recA обнаруживает ингибирование ДНК синтез и инициирует события, ведущие к протеолитическому расщеплению и инактивации репрессора пути SOS, кодируемого lexA ген.
Некоторые регулоны индуцируются определенными раздражителями окружающей среды, например, питательными веществами. ограничение или осмотический стресс. Часто опероны более чем одного регулона могут быть индуцированный, а термин «стимулон» использовался для описания набора генов, так что индуцированный. Обычно бактерии воспринимают такие условия окружающей среды двумя способами. компонентные системы. Первый компонент — это трансмембранный белок с внеклеточные и внутриклеточные домены. Его внеклеточный домен обнаруживает стимул окружающей среды, и его цитоплазматический домен передает сигнал.В второй компонент — бифункциональный цитоплазматический белок. Имеет домен-получатель который взаимодействует с модулем передатчика первого компонента, а также с эффекторный домен, контролирующий экспрессию соответствующего регулона. В модули передатчика и приемника двухкомпонентных систем регулирования от большое количество регулонов генетически связаны и имеют гомологию аминокислот. Сигнально-детектирующий и эффекторный домены белков из разных Однако регулоны различаются и определяют детектируемый сигнал и опероны, которые активируются или подавляются в ответ на этот сигнал.
Глобальное регулирование играет важную роль в физиологии патогенных бактерий. Например, Vibrio cholerae и Bordetella. pertussis выражают многие из детерминант вирулентности в управление системами передачи сигналов, относящимися к двухкомпонентной системы, описанные выше. Экспрессия белков, необходимых для инвазивного Фенотип контролируется температурой в Shigella . Yersinia enterocolitica чувствует температуры и концентрации ионов кальция и связывает эти сигналы с экспрессия генов и клеточная локализация продуктов генов, которые подходит для внутриклеточной или внеклеточной среды.В тканях хозяина концентрация свободного железа крайне низка, и большинство болезнетворных бактерий имеют системы транспорта железа с высоким сродством, которые индуцируются при низком уровне железа условия. Синтез дифтерийного токсина C. diphtheriae , токсин Shiga by Shigella dysenteriae , экзотоксин А от Pseudomonas aeruginosa и другие специфические белков у многих патогенных бактерий индуцируется в условиях ограниченного железом рост. Эти примеры показывают, как факторы окружающей среды могут регулировать экспрессия генов вирулентности у патогенных бактерий.
Генетика — Медицинская микробиология — Книжная полка NCBI
Генетическая информация в микробах
Генетический материал бактерий и плазмид — это ДНК. Бактериальные вирусы (бактериофаги или фаги) имеют ДНК или РНК в качестве генетического материала. Два основных Функции генетического материала — репликация и экспрессия. Генетический материал должен точно реплицироваться, чтобы потомство унаследовало все специфические генетические детерминанты (генотип) родительского организма. Выражение конкретных генетических материал в определенном наборе условий роста определяет наблюдаемые характеристика (фенотип) организма.Бактерии имеют мало структурных или особенности развития, которые можно легко наблюдать, но у них есть широкий спектр биохимические возможности и модели восприимчивости к противомикробным агентам или бактериофаги. Эти последние характеристики часто выбираются как наследуемые. черты, подлежащие анализу в исследованиях бактериальной генетики.
Структура нуклеиновой кислоты
Нуклеиновые кислоты — это большие полимеры, состоящие из повторяющихся нуклеотидных единиц (). Каждый нуклеотид содержит один фосфатная группа, один пентозный или дезоксипентозный сахар и один пурин или пиримидин основание.В ДНК сахар — это D-2-дезоксирибоза; в РНК сахар — это D-рибоза. В ДНК пуриновые основания представляют собой аденин (A) и гуанин (G), а пиримидиновые основания представляют собой тимин (Т) и цитозин (С). В РНК урацил (U) заменяет тимин. Химически модифицированные пуриновые и пиримидиновые основания обнаружены у некоторых бактерий и бактериофаги. Повторяющаяся структура полинуклеотидов включает чередование остатки сахара и фосфата, с фосфодиэфирными связями, связывающими 3′-гидроксильная группа одного нуклеотидного сахара к 5′-гидроксилу группа соседнего нуклеотидного сахара.Эти асимметричные фосфодиэфирные связи определяют полярность полинуклеотидной цепи. Основание пурина или пиримидина связан у 1′-углеродного атома каждого остатка сахара и выступает из повторяющийся сахарно-фосфатный остов. Двухцепочечная ДНК является спиральной, а две нити спирали антипараллельны. Двойная спираль стабилизирована водородные связи между пуриновыми и пиримидиновыми основаниями на противоположных цепях. В каждое положение, A на одной нити пары двумя водородными связями с T на противоположная нить, или G пары тремя водородными связями с C.Две нити поэтому двойные спирали ДНК комплементарны. Из-за взаимодополняемости двухцепочечная ДНК содержит эквимолярные количества пуринов (A + G) и пиримидины (T + C), где A равно T, а G равно C, но моль доля G + C в ДНК широко варьирует у разных бактерий. Информация в нуклеиновых кислотах кодируется упорядоченной последовательностью нуклеотидов. вдоль полинуклеотидной цепи, а в двухцепочечной ДНК последовательность каждого strand определяет, какой должна быть последовательность комплементарной цепи.В степень гомологии последовательностей между ДНК из разных микроорганизмов является самый строгий критерий для определения того, насколько тесно они связаны.
Рисунок 5-1
Двойная спиральная структура ДНК. На схеме представлена структура ДНК в виде спирали. лестница. Остов каждой полинуклеотидной цепи (представлен как лента) состоит из чередующихся остатков фосфата и дезоксирибозы связаны фосфодиэфиром (подробнее …)
Репликация ДНК
Во время репликации бактериального генома каждая нить в двойной спирали ДНК служит шаблоном для синтеза новой комплементарной цепи.Каждая дочь Таким образом, двухцепочечная молекула ДНК содержит одну старую полинуклеотидную цепь и одну вновь синтезированная нить. Такой тип репликации ДНК называется полуконсервативный. Репликация хромосомной ДНК у бактерий начинается с конкретный хромосомный участок, называемый источником, и продолжается двунаправленно, пока процесс завершен (). Когда бактерии делятся бинарным делением после завершения репликации ДНК, реплицированные хромосомы делятся на каждую из дочерних клеток. В области происхождения специфически и временно связываются с клеточной мембраной после того, как репликация ДНК была инициирована, что привело к модели, согласно которой мембрана прикрепление управляет разделением дочерних хромосом (модель репликона).Эти характеристики репликации ДНК во время роста бактерий соответствуют требования генетического материала к точному воспроизведению и наследуется каждой дочерней клеткой во время деления клетки.
Рисунок 5-2
Авторадиография интактной реплицирующейся хромосомы E. coli . Бактерии были радиоактивно помечены тритием тимидином для приблизительно в двух поколениях и подвергались бережному лизированию. Бактериальная ДНК затем был исследован авторадиографией. Вставка показывает репликацию бактериальный (подробнее…)
Экспрессия гена
Генетическая информация, закодированная в ДНК, выражается путем синтеза специфических РНК и белки, и информация течет от ДНК к РНК к белку. ДНК-направленный синтез РНК называется транскрипцией. Поскольку пряди двухспиральные ДНК антипараллельны и комплементарны, только одна из двух цепей ДНК может служат в качестве матрицы для синтеза конкретной молекулы мРНК. Информационные РНК (мРНК) передают информацию от ДНК, и каждая мРНК в бактериях функционирует как матрица для синтеза одного или нескольких конкретных белков.Процесс по нуклеотидная последовательность молекулы мРНК определяет первичный амино кислотная последовательность белка называется трансляцией. Рибосомы, комплексы рибосомные РНК (рРНК) и несколько рибосомных белков, транслируют каждую мРНК в соответствующую полипептидную последовательность с помощью транспортных РНК (тРНК), синтез аминоацил тРНК, факторы инициации и факторы элонгации. Все эти компоненты аппарата для синтеза белка функционируют в производство множества различных белков.Ген — это последовательность ДНК, которая кодирует белок, рРНК или молекула тРНК (продукт гена).
Генетический код определяет, как нуклеотиды в мРНК определяют аминокислоты. в полипептиде. Потому что в мРНК всего 4 разных нуклеотида. (содержащие U, A, C и G), отдельные нуклеотиды не содержат достаточно информации однозначно указать все 20 аминокислот. В динуклеотидах 16 (4 × 4) возможны расположения четырех нуклеотидов, а в Возможны 64 компоновки тринуклеотидов (4 × 4 × 4).Таким образом, требуется минимум три нуклеотида для обеспечения хотя бы одного уникального последовательность, соответствующая каждой из 20 аминокислот. В «Универсальный» генетический код, используемый большинством организмов (), представляет собой тройной код, в котором 61 из 64 возможных тринуклеотида (кодона) кодируют определенные аминокислоты, и любой из три оставшихся кодона (UAG, UAA или UGA) приводят к прекращению перевод. Кодоны, обрывающие цепь, также называются бессмысленными кодонами. потому что они не определяют никаких аминокислот. Генетический код описывается как вырожденный, потому что несколько кодонов могут использоваться для одной аминокислоты, и как неперекрывающиеся, потому что соседние кодоны не имеют общих нуклеотидов.Исключения из «универсального» кода включают использование UGA в качестве кодон триптофана у некоторых видов Mycoplasma и у митохондриальная ДНК и несколько дополнительных кодоновых различий в митохондриальной ДНК из дрожжей , Drosophila и млекопитающих. Трансляция мРНК осуществляется обычно инициируется кодоном AUG для метионина, а соседние кодоны транслируется последовательно, когда мРНК читается с 5 ‘по 3’ направление. Соответствующая полипептидная цепь собирается, начиная с ее аминоконце и продвигаясь к его карбоксиконцу.Последовательность амино кислоты в полипептиде, следовательно, колинеарны с последовательностью нуклеотидов в мРНК и соответствующем гене. Специфические ферментативные реакции участвуют в синтезе ДНК, РНК и белка, выходят за рамки этого глава.
Экспрессия генетических детерминант у бактерий включает однонаправленный поток информации от ДНК к РНК к белку. В бактериофагах либо ДНК, либо РНК. может служить генетическим материалом. При заражении бактерий РНК бактериофаги, молекулы РНК служат матрицами для репликации РНК и мРНК.Исследования ретровирусной группы вирусов животных показывают, что ДНК молекулы могут быть синтезированы из матриц РНК ферментами, обозначенными как РНК-зависимые ДНК-полимеразы (обратные транскриптазы). Этот поворот обычное направление потока генетической информации от РНК к ДНК, а не от ДНК в РНК — важный механизм передачи информации от ретровирусов. закодироваться в ДНК и стать частью геномов животных клетки.
Организация генома
Молекулы ДНК, которые реплицируются как дискретные генетические единицы в бактериях, называются репликоны.У некоторых штаммов Escherichia coli хромосома является в клетке присутствует только репликон. У других бактериальных штаммов есть дополнительные репликоны, такие как плазмиды и бактериофаги.
Хромосомная ДНК
Бактериальные геномы варьируются по размеру от примерно 0,4 × 10 9 до 8,6 × 10 9 дальтон (Да), некоторые из самых маленьких являются обязательными паразиты ( Mycoplasma ) и самые крупные, принадлежащие к бактериям способны к сложной дифференцировке, например, Myxococcus .В количество ДНК в геноме определяет максимальное количество информации, которую он может кодировать. Большинство бактерий имеют гаплоидный геном, единственную хромосому, состоящую из кольцевой двухцепочечной молекулы ДНК. Однако линейные хромосомы имеют был обнаружен у грамположительных Borrelia и Streptomyces spp., Один линейный и один круговой хромосома присутствует в грамотрицательной бактерии Agrobacterium tumefaciens . Единая хромосома общего кишечного бактерия E.coli составляет 3 × 10 9 Да (4500 пары килобаз [kbp]) размером примерно от 2 до 3 процентов от сухой массы клетки. Геном E. coli является всего около 0,1% от размера генома человека, но этого достаточно для кодирования несколько тысяч полипептидов среднего размера (40 кДа или 360 аминокислот).
Хромосома кишечной палочки имеет контурную длину приблизительно 1,35 мм, что в несколько сотен раз длиннее бактериальной клетки, но ДНК суперспиральный и плотно упакованный в бактериальный нуклеоид.Необходимое время для репликации всей хромосомы составляет около 40 минут, что составляет примерно вдвое меньше кратчайшего времени деления этой бактерии. ДНК репликация должна инициироваться так часто, как клетки делятся, чтобы быстро при выращивании бактерий новый виток хромосомной репликации начинается раньше, чем раньше. раунд завершен. При высоких темпах роста может быть четыре хромосомы. реплицируются с образованием восьми во время деления клеток, что связано с завершение раунда хромосомной репликации.Таким образом, хромосома в быстрорастущие бактерии размножаются более чем в одной точке. Репликация хромосомной ДНК бактерий сложна и включает в себя множество различных белки.
Плазмиды
Плазмиды — это репликоны, которые сохраняются как дискретные, внехромосомные генетические элементы в бактериях. Обычно они намного меньше бактериального хромосома, варьирующаяся от менее 5 до более нескольких сотен кб, хотя плазмиды размером до 2 Mbp встречаются у некоторых бактерий. Плазмиды обычно кодируют черты, которые не важны для жизнеспособности бактерий, и воспроизводятся независимо от хромосомы.Большинство плазмид имеют суперспиральную, круговую, двухцепочечные молекулы ДНК, но также были продемонстрированы линейные плазмиды у Borrelia и Streptomyces . Близко связанный или идентичные плазмиды демонстрируют несовместимость; они не могут быть стабильными сохраняется в том же бактериальном хозяине. Классификация плазмид основана на несовместимости или использования конкретных ДНК-зондов в тестах гибридизации с идентифицировать нуклеотидные последовательности, характерные для конкретной плазмиды репликоны.Некоторые гибридные плазмиды содержат более одного репликона. плазмиды кодируют функции, которые способствуют передаче плазмиды от донора бактерии к другим бактериям-реципиентам, но неконъюгативные плазмиды — нет. Конъюгативные плазмиды, которые также способствуют переносу бактериальной хромосомы из бактерии-доноры для других бактерий-реципиентов называются плазмидами фертильности, и обсуждаются ниже. Среднее количество молекул данной плазмиды на бактериальная хромосома называется числом ее копий.Большие плазмиды (> 40 пары килобаз) часто бывают конъюгативными, имеют уменьшенное количество копий (от 1 до нескольких на хромосома), код для всех функций, необходимых для их репликации, и разделяются между дочерними клетками во время деления клеток способом похож на бактериальную хромосому. Плазмиды размером менее 7,5 тысяч пар оснований обычно не сопряжены, имеют большое количество копий (обычно 10–20 на хромосому), полагаются на своего бактериального хозяина, чтобы обеспечить некоторые необходимые функции для репликации и случайным образом распределяются между дочерними клетками на разделение.
Многие плазмиды контролируют важные с медицинской точки зрения свойства патогенных бактерий, включая устойчивость к одному или нескольким антибиотикам, выработку токсинов и синтез структур клеточной поверхности, необходимых для прикрепления или колонизации. Плазмиды, определяющие устойчивость к антибиотикам, часто называют плазмидами R. (или R-факторы). Типичные токсины, кодируемые плазмидами, включают термолабильные и термостойкие энтеротоксины E. coli , эксфолиативный токсин золотистый стафилококк и столбнячный токсин Clostridium tetani .Некоторые плазмиды загадочны и не имеют узнаваемое воздействие на бактериальные клетки, в которых они находятся. Сравнение плазмиды профили — полезный метод оценки возможного родства отдельных клинические изоляты определенного вида бактерий для эпидемиологических исследований. Обсуждается роль плазмид в развитии устойчивости к антибиотикам. ниже.
Бактериофаги
Бактериофаги (бактериальные вирусы, фаги) — инфекционные агенты, которые размножаются как облигатные внутриклеточные паразиты у бактерий.Внеклеточные фаговые частицы метаболически инертны и состоят в основном из белков и нуклеиновой кислоты (ДНК или РНК, но не то и другое вместе). Белки фаговой частицы образуют защитную оболочка (капсид), окружающая плотно упакованный геном нуклеиновой кислоты. Фаг геномы различаются по размеру от примерно 2 до 200 килобаз на цепь нуклеиновой кислоты. кислоты и состоят из двухцепочечной ДНК, одноцепочечной ДНК или РНК. Фаг геномы, как и плазмиды, кодируют функции, необходимые для репликации в бактериях, но в отличие от плазмид они также кодируют белки капсида и неструктурные белки требуется для сборки фага.Несколько морфологически различных типов фага были описаны, в том числе многогранные, нитевидные и сложные. Сложный фаги имеют многогранные головы, к которым хвосты, а иногда и другие придатки (хвост пластины, хвостовые волокна и т. д.).
Один цикл роста фага показан на. Инфекция инициируется адсорбцией фага на специфических рецепторы на поверхности восприимчивых бактерий-хозяев. Капсиды остаются на поверхность клетки, а геномы ДНК или РНК проникают в клетки-мишени (проникновение).Поскольку инфекционность геномной ДНК или РНК намного меньше, чем у зрелых вируса, сразу после заражения наступает период, называемый периодом затмения. во время которого невозможно обнаружить внутриклеточный инфекционный фаг. Заражение РНК или ДНК фага реплицируются, чтобы произвести множество новых копий генома фага, и продуцируются фаг-специфические белки. Для большинства фагов сборки потомства происходит в цитоплазме, а высвобождение потомства происходит путем лизиса клеток. В напротив, нитчатые фаги образуются на клеточной оболочке и высвобождаются не убивая клетки-хозяева.Период затмения заканчивается, когда внутриклеточный появляются заразные потомства. Скрытый период — это интервал от заражения. до появления внеклеточного потомства, а период роста — это интервал от конец латентного периода, пока все фаги не станут внеклеточными. Среднее число частиц фага, продуцируемых каждой инфицированной клеткой, называемого размером взрыва, составляет характерна для каждого вируса и часто составляет от 50 до нескольких сотен. Для обсуждения строения, размножения и классификации животных. вирусы, см. главы 41 и 42.
Рисунок 5-3
Одностадийный рост бактериофага. Культура восприимчивых бактерий синхронно заражается бактериофаг добавлен в момент времени 0 при низкой множественности инфекции. Вскоре после этого неабсорбированный фаг инактивируется добавлением антифаговая антисыворотка, (подробнее …)
Фаги подразделяются на две основные группы: вирулентные и умеренные. Рост вирулентные фаги у чувствительных бактерий разрушают клетки-хозяева. Заражение восприимчивые бактерии к умеренным фагам могут иметь один из двух результатов: литический рост или лизогения.Литический рост умеренных и вирулентных бактериофагов аналогично, что приводит к образованию фагового потомства и гибели бактерий-хозяев. Лизогения — это особый тип латентной вирусной инфекции, при которой геном фага реплицируется как профаг в бактериальной клетке. У большинства лизогенных бактерий гены, необходимые для развития литического фага, не экспрессируются, и производство инфекционный фаг не встречается. Кроме того, лизогенные клетки невосприимчивы к суперинфекция вирусом, который они носят как профаги.Физическое состояние профага не является идентичным для всех вирусов умеренного климата. Например, профаг бактериофага λ в E. coli интегрирован в бактериальную хромосому в определенном месте и реплицируется как часть бактериальная хромосома, тогда как профаг бактериофага P1 у E. coli реплицируется как внехромосомная плазмида.
Рост литического фага происходит спонтанно в небольшой фракции лизогенных клеток, и несколько внеклеточных фагов присутствуют в культурах лизогенных бактерий.Для некоторых лизогенных бактерий синхронная индукция развития литических фагов встречается во всей популяции лизогенных бактерий, когда они обрабатываются агенты, которые повреждают ДНК, такие как ультрафиолет или митомицин С. Потеря профаг из лизогенной бактерии, переводя его в нелизогенное состояние и восстановление восприимчивости к инфекции первоначально присутствующим фагом как профаг, называется лечением.
Некоторые фаги умеренного пояса содержат гены бактериальных характеристик, которые не связаны с развитием литического фага или лизогенным состоянием, и экспрессия такие гены называют фаговой конверсией (или лизогенной конверсией).Примеры конверсия фагов, которая важна для микробной вирулентности, включает производство токсина дифтерии Corynebacterium diphtheriae , эритрогенный токсин Streptococcus pyogenes (группа А β-гемолитические стрептококки), ботулинический токсин от Clostridium botulinum , а шига-подобные токсины — E. coli . В В каждом из этих примеров ген, кодирующий бактериальный токсин, присутствует в умеренный геном фага. Специфичность O-антигенов в Salmonella также можно контролировать с помощью фаговой конверсии.Типирование фагов — это тестирование штаммов определенного вида бактерий на восприимчивость к специфическим бактериофагам. Паттерны восприимчивости к набор типирующих фагов предоставляет информацию о возможном родстве отдельные клинические изоляты. Такая информация особенно полезна для эпидемиологические исследования.
Мутация и отбор
Мутации — это наследуемые изменения генома. Спонтанные мутации у человека бактерии встречаются редко. Некоторые мутации вызывают изменения фенотипических характеристик; то о возникновении таких мутаций можно судить по вызываемым ими эффектам.В контрольные организмы, специфичные для микробной генетики, обозначены как штаммы дикого типа, а потомки, в геномах которых есть мутации, называются мутантами. Таким образом, мутанты характеризуются унаследованными различиями между ними и их предковые штаммы дикого типа. Вариантными формами конкретной генетической детерминанты являются: называются аллелями. Генотипические символы — строчные буквы, сокращения выделены курсивом. указать отдельные гены с надстрочным индексом (+), указывающим на дикий тип аллель. Фенотипические символы пишутся с большой буквы, а не курсивом, чтобы их можно было различить. от генотипических символов.Например, генотипический символ способности производить β-галактозидаза, необходимая для ферментации лактозы, является lacZ + и мутанты, которые не могут производить β-галактозидазы lacZ . Фенотип ферментации лактозы обозначается Lac + , а неспособность ферментировать лактозу Lac — .
Обнаружение мутантных фенотипов
Селективные и дифференциальные среды полезны для выделения бактериальных мутантов. Некоторые селективные среды позволяют расти определенным мутантам, но не позволяют штаммы дикого типа для выращивания.Редкие мутанты можно выделить с помощью таких селективных СМИ. Дифференциальные среды позволяют бактериям дикого типа и мутантным бактериям расти и формироваться. колонии, различающиеся по внешнему виду. Обнаружение редких мутантов по дифференциалу media ограничено общим количеством колоний, которые можно наблюдать. Рассматривать штамм E. coli дикого типа, чувствительный к антибиотик стрептомицин (фенотип Str s ) и может использовать лактозу в качестве единственный источник углерода (фенотип Lac + ).Спонтанно Встречающиеся мутанты Str r редки и обычно встречаются с частотами менее одной на 109 бактерий в культурах дикого типа E. coli . Тем не менее, мутанты Strr можно легко выделить, используя селективная среда, содержащая стрептомицин, поскольку Str s дикого типа бактерии погибают. Выделение лактозонегативных (фенотип Lac — ) мутанты E. coli представляют собой другая проблема. На минимальных средах с лактозой в качестве единственного источника углерода, Lac + штаммы дикого типа будут расти, но Мутанты Lac — не могут расти.На дифференциальных средах, таких как МакКонки-лактозный агар или эозин-метиленовый синий-лактозный агар, Lac + дикого типа и мутантные штаммы Lac — E. coli можно отличить по цвету, но самопроизвольно Мутанты Lac — слишком редки, чтобы их легко выделить. Селективная среда для мутантов Lac — E. coli можно получить путем включения химических аналогов лактозы, которые превращаются в токсичные метаболиты бактериями Lac + , но не мутантами Lac —.Мутанты Lac — затем могут расти на такой среде, но бактерии дикого типа Lac + убиты.
Мутации, инактивирующие основные гены гаплоидных организмов, обычно летальные, но такие потенциально смертельные мутации часто можно изучить, если их экспрессия контролируется манипулированием экспериментальными условиями. Для Например, мутация, которая увеличивает термолабильность важного гена продукт может предотвратить рост бактерий при 42 ° C, хотя мутант бактерия все еще может расти при 25 ° C.И наоборот, чувствительные к холоду мутанты выражают мутантный фенотип при низкой температуре, но не при высокой температуре. Чувствительные к температуре и холоду мутации являются примерами условных мутации, как и подавляемые мутации, описанные далее в этой главе. А условный летальный фенотип указывает на то, что мутантный ген важен для жизнеспособность.
Спонтанные и индуцированные мутации
Скорость мутации у бактерий определяется точностью репликации ДНК, возникновение повреждения ДНК и эффективность механизмов восстановления поврежденной ДНК.
Для конкретного бактериального штамма в определенных условиях роста мутация скорость для любого конкретного гена постоянна и выражается как вероятность мутации на деление клетки. В популяции бактерий, выросших из небольшого inoculum, доля мутантов обычно прогрессивно увеличивается по мере увеличения размера бактериальной популяции увеличивается.
Мутации у бактерий могут происходить спонтанно и независимо от для их обнаружения используются экспериментальные методы. Этот принцип был впервые продемонстрирован тестом на колебания ().В количество устойчивых к фагам мутантов E. coli в повторностях культуры, выращенные из небольших инокул, были измерены и сравнены с культурами, выращенными в несколько образцов, взятых из одной культуры. Если мутации по устойчивости к фагам произошла только после воздействия фага, вариабельность количества мутантов между культурами должны быть одинаковыми в обоих наборах условий. Напротив, если устойчивые к фагам мутанты возникли спонтанно до воздействия от бактерий к фагу, количество мутантов должно быть более вариабельным в самостоятельно выращенные культуры, поскольку различия в размерах бактерий популяции, когда появился первый мутант, будет способствовать наблюдаемому изменчивость.Данные показали, что мутации устойчивости к фагам в E. coli возник спонтанно с постоянной вероятностью на деление клетки.
Рисунок 5-4
Испытание на колебания. Различия в количестве колоний устойчивых к фагам мутантов в повторные образцы из одной субкультуры были небольшими и отражали только ожидаемые колебания из-за ошибок выборки. Напротив, количество устойчивых к фагам колоний (подробнее …)
Посев на реплики подтвердил, что мутации у бактерий могут возникать спонтанно, без воздействия на бактерии селективных агентов ().Для покрытия реплик: плоский стерильный вельветовый поверхность используется для сбора инокулята с поверхности эталонной чашки с агаром и перенесите образцы в другие чашки с агаром. Таким образом, образцы Популяция бактерий с мастер-планшета переносится на реплики не искажая их пространственного расположения. Если реплики содержат селективной среды и мастер-планшетов нет, положения отобранных мутантов можно отметить колонии на чашках-репликах и бактерии, которые не подвергались воздействию к условиям отбора могут быть изолированы с тех же позиций на мастер пластина.Мутанты E. coli , устойчивые к бактериофагу Т1 или стрептомицину были выделены таким образом, не подвергая бактерии дикого типа к бактериофагу или антибиотику.
Рисунок 5-5
Выявление ранее существовавших бактериальных мутантов по реплике покрытие. Мастер-планшет был сильно засеян образцом из чистых культур чувствительные к фагам бактерии. После инкубации бактерии от хозяина планшеты были перенесены путем посева реплик в дубликаты (подробнее …)
На частоту мутаций влияют как экологические, так и генетические факторы.Контакт с бактерий в мутагенные агенты вызывает увеличение скорости мутаций, иногда на на несколько порядков. Многие химические и физические агенты, включая рентгеновские лучи и ультрафиолет, обладают мутагенной активностью. Канцерогенные химические вещества для животных часто являются мутагенными для бактерий или могут быть преобразованы животными ткани на метаболиты, которые являются мутагенными для бактерий. Стандартизированные тесты для мутагенность бактерий используются в качестве скрининговых процедур для выявления агенты окружающей среды, которые могут быть канцерогенными для человека.Гены-мутаторы в бактерии вызывают увеличение частоты спонтанных мутаций для самых разных другие гены. Экспрессия этих генов, вызванная повреждением ДНК (см. SOS-ответ позже), позволяет восстанавливать повреждения ДНК, которые в противном случае были бы летальными, но за счет подверженный ошибкам механизм, который увеличивает скорость мутации. В целом скорость мутации — вероятность того, что мутация произойдет где-то в бактериальный геном на деление клетки — относительно постоянен для множество организмов с геномами разного размера и, по-видимому, важный фактор в определении пригодности бактериального штамма к выживанию в природе.Большинство мутаций вредны, и существует риск неблагоприятных мутаций для отдельные бактерии должны быть сбалансированы с положительным значением изменчивости, поскольку механизм адаптации бактериальных популяций к изменяющимся условиям окружающей среды. условия.
Молекулярная основа мутаций
Мутации классифицируются на основе структурных изменений, происходящих в ДНК (). Некоторые мутации локализованы в коротких сегментах ДНК (например, при нуклеотидных заменах, микроделеции и микровставки).Другие мутации затрагивают большие области ДНК и включают делеции, вставки или перестройки сегментов ДНК.
Когда нуклеотидная замена происходит в области ДНК, которая кодирует полипептид, один из трех нуклеотидов в одном кодоне соответствующая молекула мРНК будет изменена. Тихие мутации не меняют структура или функция полипептида, потому что один кодон в мРНК заменен на другой для той же аминокислоты. Другие замены вызывают замену одной аминокислоты. заменен другим в конкретном положении в полипептиде соответствующий измененному кодону.Мутации, приводящие к замене одного аминокислота для другой в полипептидной цепи называется миссенс-мутациями. Влияние аминокислотных замен на функцию полипептидного гена продукт различается и зависит от местоположения и идентичности аминокислоты замена. Мутантные полипептиды, содержащие замены аминокислот, обычно имеют общие антигенные детерминанты с полипептидом дикого типа и часто имеют некоторые остаточная биологическая активность. Мутации, приводящие к замене аминокислоты кислотный кодон с терминирующим кодоном называют нонсенс-мутациями.Это результаты в продукции амино-концевого фрагмента нормального полипептида, когда мутантная мРНК транслируется. Бессмысленные мутации часто приводят к полной потере активность генного продукта.
Из-за триплетной природы генетического кода последствия мутаций вызванные вставками или удалениями небольшого количества нуклеотидов (микровставки, микроделеции) зависят как от количества, так и от последовательности вовлеченные нуклеотиды. Удаление или добавление кратных трех нуклеотидов пары не влияют на рамку считывания, но вызывают удаление или добавление соответствующее количество аминокислот в одном сайте полипептида.Если новый кодон обрыва цепи вводится, преждевременное обрыв цепи происходит в пределах полипептид. Напротив, добавление или удаление других номеров пары нуклеотидов изменяют рамку считывания для всего сегмента мРНК от мутация дистального конца гена. Следовательно, мутации сдвига рамки считывания может вызвать резкие изменения в структуре и активности полипептида генные продукты, и их часто классифицируют как бессмысленные мутации.
Тесты комплементации
Чтобы определить, находятся ли мутации в одном и том же гене или в разных генах, тесты комплементации проводятся с частично диплоидными бактериальными штаммами ().Две копии региона бактериальной хромосомы, несущей мутацию, присутствуют в том же бактерия, причем каждая копия содержит разные мутации (мутации находятся в транс расположение). Фенотип дикого типа указывает на то, что мутации разные гены. Это явление называется комплементацией. Если мутант фенотипа, необходимо провести контрольный эксперимент с мутации в цис-расположении, чтобы исключить возможность того, что дикий тип аллели не могут нормально экспрессироваться в частично диплоидном штамме бактерий.Изначально тесты комплементации назывались «цис-транс». тесты, а термин цистрон иногда используется как синоним гена. Дополнительные тесты могут быть выполнены и интерпретированы, даже если биохимические функции продуктов гена неизвестны.
Рисунок 5-6
Комплементация — это метод проверки функционального гена товары. Два мутанта со схожими фенотипами (неспособность конвертировать субстрат X к продукту Z). Мутации в этих штаммах обозначены a и b, соответственно, и аллели дикого типа (подробнее…)
В качестве примера рассмотрим использование теста дополнения для характеристики двух независимо полученные мутанты Lac — E. coli . Биохимический путь утилизации лактозы требует β-галактозид пермеаза (генотипический символ lac Y) для транспортировки лактоза в бактериальную клетку и β-галактозидазу (генотипический символ lacZ ) для превращения лактозы в D-глюкозу и D-галактозу. Мутанты без β-галактозид пермеазы или β-галактозидазы не могут используйте лактозу для роста.Если мутации в обоих Lac — мутанты инактивировали тот же белок (например, β-галактозид), затем частичный диплоидный штамм, содержащий генов lacZ от обоих мутанты в транс-структуре не могут использовать лактозу. В напротив, если бы генотипы двух мутантов были lacZ + lacY и lacZ lacY + , частично диплоидная бактерия будет производить активную β-галактозидазу из lacZ + определитель и активный β-галактозид пермеаза из лаковый + определитель.Дополнение произойдет, и частично диплоидный штамм будет использовать лактозу.
Реверсия и подавление
Мутации, которые преобразуют фенотип из дикого типа в мутантный, называются прямыми мутации и мутации, которые изменяют фенотип с мутантного обратно на дикий тип называются обратными мутациями (реверсиями). Бактериальные штаммы, содержащие обратные мутации называются ревертантами. Анализ мутаций, вызывающих фенотипическая реверсия дает полезную информацию. Обратные мутации, восстанавливающие точные нуклеотидные последовательности ДНК дикого типа являются истинными реверсиями.Истинный ревертанты идентичны штаммам дикого типа как генотипически, так и фенотипически. Обратные мутации, которые не восстанавливают точный нуклеотид Последовательности ДНК дикого типа называются супрессорными мутациями (супрессорами). Некоторые ревертанты, несущие супрессорные мутации, фенотипически неотличимы от штаммов дикого типа. Другие ревертанты, называемые псевдоревертанты, фенотипически отличимые от штаммов дикого типа, например, из-за тонких различий в характеристиках ферментативного восстановленная активность (например, удельная активность, субстрат специфичность, кинетические константы или восприимчивость к термическому или химическому инактивация).Распознавание псевдоревертантных фенотипов предполагает наличие супрессорных мутаций.
Супрессорные мутации могут быть внутригенными или экстрагенными. Внутригенные супрессоры расположены в том же гене, что и подавляемые ими прямые мутации. В возможные места и характер внутригенных супрессоров определяются исходная прямая мутация и отношения между первичной структурой продукта гена и его биологической активности. Экстрагенные супрессоры находятся в разных генах от мутаций, эффекты которых они подавляют.В способность экстрагенных супрессоров подавлять множество независимых мутаций можно протестировать. Некоторые экстрагенные супрессоры специфичны для определенных генов, некоторые специфичны для определенных кодонов, а некоторые имеют другую специфичность узоры. Экстрагенные супрессоры, которые меняют фенотипические эффекты кодоны, завершающие цепь, были хорошо охарактеризованы, и было обнаружено, что они изменяют структура специфических тРНК. Конкретная супрессорная тРНК может разрешать определенные кодон обрыва цепи, который будет транслироваться, что приведет к включению специфической аминокислоты в формирующийся полипептид в положении, соответствующем к кодону обрыва цепи.У бактерии, имеющей оканчивающуюся цепь мутация и соответствующий экстрагенный супрессор, трансляция мРНК содержащий мутантный кодон, следовательно, может привести к образованию полноразмерного полипептид. Биологическая активность полноразмерного полипептида, образованного как Последствия подавления зависят как от количества вырабатываемого белка, так и от функциональные последствия замены конкретной аминокислоты, определяемые супрессорная тРНК.
Обмен генетической информацией
Биологическое значение сексуальности у микроорганизмов заключается в увеличении вероятность того, что редкие независимые мутации произойдут вместе в одном микробе и подвергнуться естественному отбору.Генетические взаимодействия между микробами позволяют их геномы эволюционируют гораздо быстрее, чем за счет одной лишь мутации. Представитель явления, имеющие медицинское значение, которые включают обмен генетической информацией или геномные перестройки включают быстрое появление и распространение антибиотика. плазмиды устойчивости, вариации жгутиковых фаз у сальмонелл и антигенные изменение поверхностных антигенов у Neisseria и Borrelia.
Половые процессы у бактерий включают передачу генетической информации от донора к реципиент и приводит либо к замене аллелей донора на аллели реципиента или добавление донорских генетических элементов в геном реципиента.Трансформация, трансдукция и конъюгация — это половые процессы, в которых используются разные механизмы для ввести донорскую ДНК в реципиентные бактерии (). Поскольку донорская ДНК не может сохраняться в бактерии-реципиенте, если это часть репликона, рекомбинация между геномами донора и реципиента часто требуется для получения стабильного гибридного потомства. Наиболее вероятна рекомбинация когда донорская и реципиентная бактерии происходят от одного или близкородственного происхождения разновидность.
Рисунок 5-7
Обмен генетической информацией в бактериях.Трансформация, трансдукция и конъюгация различаются способами введение ДНК из клетки-донора в клетку-реципиент. А) В трансформация, отбираются фрагменты ДНК, освобожденные от донорских бактерий вверх по компетентному (подробнее …)
Для того, чтобы рекомбинант был обнаружен, его фенотип должен отличаться от обоих родительских фенотипы. Перед рекомбинантным фенотипом может потребоваться рост или деление клеток. выражается. Задержка экспрессии рекомбинантного фенотипа до гаплоида сегрегация рекомбинантного генома называется задержкой сегрегации и задержкой до тех пор, пока синтез продуктов, кодируемых донорскими генами, называется фенотипическим отставанием.Тестирование на сцепление (неслучайная перегруппировка родительских аллелей в рекомбинантных потомство) возможно, когда родительские бактерии имеют разные аллели для нескольких гены. Донорский аллель невыбранного гена с большей вероятностью присутствует в рекомбинантов, если он связан с выбранным донорским геном, чем если он не связан с выбранный донорский ген. Количественный анализ связей позволяет построить генетические карты. Геном E. coli является кольцевым (), что определяется как генетическими сцепление и прямой биохимический анализ хромосомной ДНК, и генетическая карта колинеарно с физической картой хромосомной ДНК.Генетическое и физическое картирование также используются для анализа внехромосомных репликонов, таких как бактериофаги и плазмиды.
Рисунок 5-8
Круговая генетическая карта E. coli . Позиции репрезентативных генов указаны на внутреннем круге. Расстояния между генами калибруются в минутах, в зависимости от времени. требуется для переноса при конъюгации. Положение треонина (th) локус произвольно (подробнее …)
Многие бактерии имеют системы рестрикции-модификации (РМ), состоящие из модифицирующих ферменты, которые метилируют остатки аденина или цитозина в определенных последовательностях в их собственная ДНК и соответствующие эндонуклеазы рестрикции, которые расщепляют чужеродную ДНК, которая не несет специфической модификации в тех же целевых последовательностях.Некоторые рестрикционные ферменты будут расщеплять только ДНК, метилированную в определенных последовательности. Эти системы ограничения, которые могли развиться для защиты бактерий против вторжения фагов или плазмид, являются важным барьером для генетических обмены между различными бактериальными штаммами или видами. Недавние данные свидетельствуют о что плазмидные системы RM могут быть способом для плазмиды обеспечить ее перенос в штамм-хозяин, поскольку клетки, теряющие плазмиду (и соответствующие защитные ген метилазы) убиваются под действием более стабильного рестрикционного фермента, который атакует недавно реплицированную, но неизмененную хромосомную ДНК.
Трансформация
При трансформации захватываются фрагменты ДНК, высвобожденные из донорских бактерий. непосредственно из внеклеточной среды бактериями-реципиентами. Рекомбинация происходит между отдельными молекулами трансформирующей ДНК и хромосомами реципиентные бактерии. Чтобы быть активными в трансформации, молекулы ДНК должны находиться на длиной не менее 500 нуклеотидов, и трансформирующая активность быстро разрушается обработкой ДНК дезоксирибонуклеазой. Молекулы трансформирующей ДНК соответствуют к очень маленьким фрагментам бактериальной хромосомы.Котрансформация генов поэтому маловероятно, если они не связаны настолько тесно, что могут быть кодируется на одном фрагменте ДНК. Трансформация была обнаружена в Streptococcus pneumoniae и встречается у других бактериальных родов, в том числе Haemophilus , Neisseria , Bacillus и Staphylococcus . Способность бактерий для поглощения внеклеточной ДНК и трансформации, называемой компетентность зависит от физиологического состояния бактерий.Многие бактерии которые обычно некомпетентны, могут быть использованы для анализа ДНК в лаборатории манипуляции, такие как кальциевый шок или воздействие высоковольтного электрического пульс (электропорация). У некоторых бактерий (включая Haemophilus и Neisseria) Поглощение ДНК зависит от наличия определенных олигонуклеотидных последовательностей в трансформирует ДНК, но у других (включая Streptococcus pneumoniae) захват ДНК не зависит от последовательности. Компетентные бактерии также могут занимать неповрежденные ДНК бактериофага (трансфекция) или плазмидная ДНК, которая затем может реплицироваться как внехромосомные генетические элементы в бактериях-реципиентах.Напротив, фрагмент хромосомной ДНК от бактерии-донора обычно не может реплицироваться в бактерия-реципиент, если она не станет частью репликона путем рекомбинации. Исторически сложилось так, что характеристика «трансформирующего принципа» из S. pneumoniae при условии, что первое прямое доказательство наличия ДНК генетический материал.
Трансдукция
При трансдукции бактериофаги действуют как векторы для введения ДНК от донора бактерии в бактерии-реципиенты путем инфицирования. Для некоторых фагов, называемых генерализованные трансдуцирующие фаги, небольшая фракция вирионов, продуцируемых во время литический рост аберрантный и содержит случайный фрагмент бактериального генома вместо фаговой ДНК.Каждый индивидуальный трансдуцирующий фаг несет свой набор тесно связанных генов, представляющих небольшой сегмент бактериального генома. Трансдукция, опосредованная популяциями таких фагов, называется генерализованной. трансдукция, потому что каждая часть бактериального генома имеет приблизительно такая же вероятность передачи от донора к бактериям-реципиенту. Когда генерализованный трансдуцирующий фаг заражает реципиентную клетку, экспрессия передаются донорские гены. Абортивная трансдукция относится к временным экспрессия одного или нескольких донорских генов без образования рекомбинантного потомства, тогда как полная трансдукция характеризуется образованием стабильных рекомбинанты, которые наследуют донорские гены и сохраняют способность их экспрессировать.В прерванная трансдукция донорский фрагмент ДНК не реплицируется, и среди потомство исходного трансдуктанта только одна бактерия содержит донорскую ДНК фрагмент. У всех остальных потомков продукты донорских генов постепенно становятся разбавляется после каждого поколения роста бактерий до тех пор, пока донорский фенотип не сможет больше не выражаться. На селективной среде, на которой только бактерии с донорский фенотип может расти, абортивные трансдуктанты производят крошечные колонии, которые легко отличить от колоний стабильных трансдуктантов.Частота абортивной трансдукции обычно на один-два порядка больше чем частота генерализованной трансдукции, что указывает на то, что большинство клеток инфицированные генерализованными трансдуцирующими фагами, не продуцируют рекомбинантные потомство.
Специализированная трансдукция отличается от обобщенной трансдукции несколькими способами. Это опосредуется только определенными умеренными фагами и лишь несколькими конкретными донорами. гены могут передаваться бактериям-реципиентам. Специализированные трансдуцирующие фаги образуются только тогда, когда лизогенные донорские бактерии входят в литический цикл и высвобождают фаговое потомство.Специализированные фаги-трансдукторы — это редкие рекомбинанты, которые не имеют части нормального генома фага и содержат часть бактериального хромосома расположена рядом с местом прикрепления профага. Многие специализированные трансдуцирующие фаги являются дефектными и не могут завершить литический цикл фага рост в инфицированных клетках, если не присутствуют фаги-помощники, функции фага. Специализированная трансдукция является результатом лизогенизации бактерия-реципиент специализированным трансдуцирующим фагом и экспрессией донорские гены.Конверсия фагов и специализированная трансдукция имеют много сходства, но происхождение генов конверсии в умеренном климате фаги неизвестны.
Конъюгация
При конъюгации прямой контакт между донорскими и реципиентными бактериями приводит к установление цитоплазматического мостика между ними и перенос части или всего донорского генома реципиенту. Донорская способность определяется конкретными конъюгативные плазмиды называются плазмидами фертильности или половыми плазмидами.
Плазмида F (также называемая фактором F) E.coli — прототип для плазмид фертильности грамотрицательных бактерий. Штаммы E. coli с внехромосомной плазмидой F называются F + и функционируют как доноры, тогда как штаммы, лишенные Плазмиды F представляют собой F — и ведут себя как реципиенты. Спряжение функции плазмиды F определяются кластером из не менее 25 переносов ( tra ) гены, определяющие экспрессию F-пилей, синтез и перенос ДНК во время спаривания, вмешательство в способность F + бактерии для выполнения и других функций.Каждая бактерия F + имеет от 1 до 3 пилей F, которые связываются с определенным белок внешней мембраны (продукт гена ompA) на бактериях-реципиентах, чтобы инициировать вязка. Образуется межклеточный цитоплазматический мостик, и одна нить F плазмидная ДНК передается от донора к реципиенту, начиная с уникального источника и прогрессирует в направлении от 5 ‘до 3’. Переданный цепь превращается в кольцевую двухцепочечную плазмидную ДНК F у реципиента бактерии, и у донора синтезируется новая цепь для замены перенесенная прядь.Обе экзонъюгантные бактерии — F + , поэтому плазмида F может распространяться инфекцией среди генетически совместимых популяции бактерий. Помимо роли F-пилей в конъюгации, они также функционируют как рецепторы для донор-специфичных (специфичных для мужчин) фагов.
Плазмида F в E. coli может существовать как внехромосомная генетический элемент или быть интегрированным в бактериальную хромосому (). Поскольку плазмида F и Бактериальные хромосомы представляют собой обе кольцевые молекулы ДНК, взаимная рекомбинация между ними образуется более крупный круг ДНК, состоящий из вставленной плазмидной ДНК F линейно в хромосому. E. coli содержит несколько копий нескольких различных генетических элементов, называемых последовательностями вставки (см. транспозоны для более подробной информации), в различных местах в его хромосоме и в Плазмида F. Гомологичная рекомбинация между последовательностями вставки в хромосома и плазмида F приводит к преимущественной интеграции плазмиды F на хромосомных участках, где расположены последовательности вставки. Хромосомный сайты, где обнаруживаются последовательности вставки, различаются, однако, среди штаммов E.coli .
Рисунок 5-9
Роль плазмиды F в определении состояний донора и реципиента E. coli . Плазмида F представляет собой специфические конъюгативные плазмиды, которые контролировать донорскую способность E. coli . F- штаммы отсутствуют плазмиды F и являются генетическими реципиентами. F + штаммы гавани (подробнее …)
Штамм E. coli с интегрированной плазмидой F сохраняет свою способность выступать донором в супружеских отношениях. Поскольку донорский штамм с интегрированные факторы F могут передавать хромосомные гены реципиентам с высоким КПД, они называются Hfr ( H igh f Requency r ecombination) штаммов.Перенос одноцепочечной ДНК от донора Hfr к реципиенту начинается от источника внутри плазмиды F и происходит, как описано выше, за исключением того, что перенесенная ДНК является гибридной. репликон, состоящий из плазмиды F, интегрированной в бактериальную хромосому. Перенос всего этого репликона, включая бактериальную хромосому, требует примерно 100 минут. Идентичность первого хромосомного гена, который будет переносится и полярность хромосомного переноса определяется сайтом интеграции плазмиды F и ее ориентации относительно бактериальная хромосома.Потому что спаривающиеся бактерии обычно отделяются спонтанно до того, как вся хромосома будет перенесена, конъюгация обычно переносится только фрагмент донорской хромосомы в реципиент. Вероятность того, что донорский ген войдет в бактерию-реципиент во время уменьшения конъюгации, следовательно, как его расстояние от начала F (и, следовательно, время его передача) увеличивается. Спаривающиеся клетки также могут быть разделены экспериментально с помощью подвергая их сильным сдвигающим усилиям в механическом блендере; это называется прерванной вязкой.Формирование рекомбинантного потомства требует рекомбинация между перенесенной донорской ДНК и геномом реципиента бактерия. Анализ потомства от вязок, прерванных после разных интервалы демонстрируют, какие хромосомные гены передаются первыми конкретных донорских штаммов, последовательное время входа для генов, которые передаются впоследствии, и все более низкая вероятность того, что гены перенесенные позже появятся в рекомбинантном потомстве. Круглость генетическая карта E.coli был первоначально выведен из перекрывающиеся, циклически переставленные группы связанных генов, которые были перенесены рано отдельными донорскими штаммами, в которых фактор F был интегрирован на разные хромосомные места.
При скрещивании бактерий F + и F — , только плазмида F передается реципиентам с высокой эффективностью. Хромосомные гены передаются с очень низкой эффективностью, и это спонтанные мутанты Hfr в популяциях F + , которые опосредуют передача донорских хромосомных генов.В вязках между Hfr и F — штаммы , сегмент плазмиды F, содержащий tra область переносится последней, после всего бактериального хромосома перенесена. Большинство рекомбинантов от вязок между Hfr и F — клетки не могут наследовать весь набор плазмиды F. гены и фенотипически являются F —. В вязках между F + и F — штаммы , плазмида F быстро распространяется среди бактериальной популяции, и большинство рекомбинантов являются Факс + .
Интегрированные плазмиды F в штаммах Hfr иногда могут быть вырезаны из бактериального хромосома. Если вырезание полностью обращает процесс интегрирования, F + клетки производятся. Однако в редких случаях иссечение происходит путем рекомбинации с участием инсерционных последовательностей или других генов на бактериальном хромосомы, расположенные на некотором расстоянии от исходного сайта интеграции. В таких случаях сегменты бактериальной хромосомы могут включаться в гибридные плазмиды F, которые называют плазмидами F ‘(см.).Подобными процессами сегменты бактериального хромосома иногда может быть включена в плазмиды R с образованием гибридных R ‘плазмиды. Конъюгативные плазмиды R ‘могут функционировать как плазмиды фертильности, потому что они могут интегрироваться в бактериальную хромосому путем гомологичной рекомбинации и опосредуют передачу хромосомных генов во время спаривания с бактериями-реципиентами. F ‘ плазмиды, R ‘плазмиды, специализированные трансдуцирующие фаги и рекомбинантные плазмиды или фаги, сконструированные путем клонирования генов (описанные ниже), представляют собой гибридные репликоны которые могут включать сегменты бактериальной хромосомы.Следовательно, любой из этих генетические элементы могут быть использованы для создания частично диплоидных бактериальных штаммы, необходимые для тестов на комплементарность и других целей.
Конъюгация также происходит у грамположительных бактерий. Грамположительные донорские бактерии вырабатывают адгезины, которые заставляют их агрегироваться с реципиентными клетками, но половые пили не участвуют. У некоторых видов Streptococcus , реципиент бактерии производят внеклеточные половые феромоны, которые вызывают изменение фенотипа донора. экспрессироваться бактериями, несущими соответствующую конъюгативную плазмиду, и конъюгативная плазмида не дает донорским клеткам продуцировать соответствующие феромон.
Рекомбинация
Рекомбинация включает разрыв и соединение родительских молекул ДНК с образованием гибридные, рекомбинантные молекулы. Было обнаружено несколько различных видов рекомбинации. идентифицированы, которые зависят от различных характеристик участвующих геномов и требуют активности различных генных продуктов. Специфические ферменты, которые действуют на ДНК (например, экзонуклеазы, эндонуклеазы, полимеразы, лигазы) участвуют в рекомбинации. Подробное обсуждение биохимических событий в рекомбинации выходит за рамки данной главы.
Обобщенная рекомбинация включает молекулы донорной и реципиентной ДНК, которые имеют гомологичные нуклеотидные последовательности. Взаимные обмены могут происходить между любыми гомологичные сайты-доноры и реципиенты. В E. coli продукт гена recA важен для генерализованной рекомбинации, но и другие генные продукты тоже участвуют.
Сайт-специфическая рекомбинация включает взаимные обмены только между определенными сайты в молекулах донорской и реципиентной ДНК. Ген recA продукт не требуется для сайт-специфической рекомбинации.Интеграция умеренный бактериофаг l в хромосому E. coli хорошо изученный пример сайт-специфической рекомбинации (). Конкретные сайты вложений (att) на Хромосома E. coli и ДНК фага λ имеют общие основная последовательность из 15 нуклеотидов, внутри которой происходит реципрокная рекомбинация, фланкированы соседними последовательностями, не гомологичными фаговым и бактериальным геномы. В фаге λ продукт гена int (интеграза) требуется для специфичного для сайта интеграционного события в лизогенизация; продукты int и Гены xis (эксцизионаза) необходимы для дополнительное сайт-специфическое иссечение, которое происходит во время индукции литического развитие фага в лизогенных клетках.
Рисунок 5-10
Интеграция и удаление бактериофага λ примеры сайт-специфической рекомбинации. λ ДНК показана тонкими линиями, а хромосомная ДНК — толстыми линиями. линий. Места прикрепления (атт) представляют собой закрытые ящики для бактериального хромосома и открытые боксы (подробнее …)
Незаконная рекомбинация — это термин, используемый для описания негомологичных, аберрантных рекомбинационные события, такие как те, которые участвуют в формировании специализированных трансдуцирующие фаги. Механизмы нелегитимной рекомбинации: неизвестный.
Транспозоны
Транспозоны — это сегменты ДНК, которые могут перемещаться из одного участка молекулы ДНК в другие сайты-мишени в той же или другой молекуле ДНК. Процесс называется транспозицией и происходит с помощью механизма, который не зависит от обобщенная рекомбинация. Транспозоны являются важными генетическими элементами, потому что они вызывают мутации, опосредуют геномные перестройки, функционируют как переносимые области генетической гомологии, приобретают новые гены и вносят свой вклад в их распространение среди бактериальных популяций.Часто введение транспозона прерывает линейную последовательность гена и инактивирует его. Транспозоны имеют важную роль в возникновении делеций, дупликаций и инверсий сегментов ДНК, как а также слияния репликонов. Транспозоны не являются самовоспроизводящимися генетическими элементы, однако, и они должны интегрироваться в другие репликоны для поддержания стабильно в бактериальных геномах.
Большинство транспозонов имеют ряд общих черт. Каждый транспозон кодирует функции, необходимые для его транспозиции, в том числе фермент транспозазы, который взаимодействует со специфическими последовательностями на концах транспозона.В течение при транспозиции короткая последовательность целевой ДНК дублируется, и транспозон вставлен между непосредственно повторяющимися целевыми последовательностями. Длина этого короткая дупликация варьирует, но характерна для каждого транспозона. В Предполагается, что дупликация включает асимметричное расщепление ДНК в мишени. сайт с последующим синтезом новых комплементарных цепей, соответствующих область между сайтами расщепления. Некоторые транспозоны вставляются практически в любые целевая последовательность, тогда как другие имеют относительно строгую целевую специфичность.Различают два типа транспозиции. Удаление транспозона из донорский сайт с последующей его вставкой в целевой сайт называется нерепликативным. транспозиция. Если транспозон на донорском сайте реплицируется и копия вставлен в целевой сайт, однако процесс называется репликативным транспозиция. Процесс репликативной транспозиции может включать образование коинтеграция, одиночная кольцевая молекула ДНК, состоящая из двух репликонов, соединенных с копиями транспозона в чередующейся последовательности.Резолюция коинтеграция в составляющие его репликоны часто достигается за счет кодируемая транспозоном резольваза, которая катализирует сайт-специфическую рекомбинацию между транспозоны. Обобщенная рекомбинация между гомологичными транспозонами может также приводят к образованию или расслоению коинтегратов. Транспозиция отличается от сайт-специфической рекомбинации путем дублирования сегмента целевой последовательности и с использованием множества различных целевых последовательностей для одного донора последовательность.
Большинство транспозонов в бактериях можно разделить на три основных класса ().Последовательности вставки и связанные составные транспозоны составляют первый класс. Последовательности вставки самые простые в структуре и кодируют только функции, необходимые для транспонирования. Известный Последовательности вставки варьируются по длине примерно от 780 до 1500 нуклеотидов. пары, имеют короткие (15-25 пар оснований) перевернутые повторы на концах и не являются тесно связаны друг с другом. ДНК между инвертированными концевыми повторами содержит один (или редко два) гена транспозазы и не кодирует резольвазу.Сложные транспозоны различаются по длине от примерно 2000 до более чем 40000. пары нуклеотидов и содержат последовательности вставки (или близкородственные последовательности) на каждом конце, обычно в виде перевернутых повторов. Весь сложный элемент может транспонировать как единое целое. ДНК между концевыми вставками последовательностей комплекса транспозоны кодируют несколько функций, которые не важны для транспозиции. У важных с медицинской точки зрения бактерий гены, определяющие приверженность к лечению. антигенов, токсинов или других факторов вирулентности, или указать устойчивость к одному или больше антибиотиков, часто находятся в сложных транспозонах.Известные примеры сложных транспозонов Tn 5 и Тн 10 , определяющие устойчивость к канамицину и тетрациклин соответственно. Сложные транспозоны, вероятно, эволюционируют транспозиция гомологичных последовательностей вставок в близлежащие сайты в ДНК молекула.
Рисунок 5-11
Характеристики репрезентативных транспозонов (жирные линии) интегрированы в бактериальную хромосому (тонкие морщинки). Транспозоны являются важными генетическими элементами, потому что они вызывают мутации, опосредуют геномные перестройки, функционируют как переносимые области генетического (подробнее…)
Второй класс транспозонов состоит из высокогомологичных ТН Семейство . Эти транспозоны имеют более длинные (от 35 до 40 пара оснований) концевые инвертированные повторы, чем описанные сложные транспозоны выше, но в них отсутствуют концевые инсерционные последовательности. Все члены семьи кодируют функции транспозазы и резольвазы. Хорошо известные примеры из Tn Семейство транспозонов включает ампициллин резистентность транспозона Tn 3 и Tn 1000 (гамма-дельта транспозон), обнаруженный в F плазмида.Семья TnA занимает важное место в истории медицины. микробиология. Развитие высокой устойчивости к ампициллину у Haemophilus influenzae и Neisseria gonorrhoeae в 1970-е годы, что сильно ограничивало полезность ампициллина для лечения гонореи и гемофильной инфекции в регионах там, где такие штаммы стали преобладать, было вызвано распространением ампициллина детерминанты устойчивости из транспозонов Tn A в плазмидах Enterobacteriaceae к плазмидам в Haemophilus и Нейссерия .
Третий класс транспозонов состоит из бактериофага Mu и родственных ему умеренных фаги. Весь геном фага функционирует как транспозон, и репликация ДНК фага во время вегетативного роста происходит путем репликативной транспозиции. Интеграция профагов может происходить во многих разных участках бактериального хромосома и часто вызывает мутации. По этой причине Mu и родственные фаги иногда их называют фагами-мутаторами.
Четвертый класс транспозонов, обнаруженный у грамположительных бактерий и представлен Тн 917 , состоит из конъюгативных транспозонов которые полностью отличаются от транспозонов, описанных выше.В конъюгативный транспозон не создает дупликации целевой последовательности в которую он вставляется, а у грамположительных бактерий штамм-хозяин, несущий транспозон может действовать как супружеский донор. Бактерии-реципиенты не нужно близкородственен донорской бактерии. Транспозон иссекают из хромосома донора и передается путем конъюгации реципиенту, где он случайным образом интегрируется в хромосому. Tn 917 кодирует устойчивость к тетрациклину, но другие более крупные конъюгативные транспозоны могут кодировать дополнительная устойчивость к антибиотикам.Конъюгативные транспозоны, по-видимому, являются основным причина распространения устойчивости к антибиотикам у грамположительных бактерий.
Некоторые роли транспозонов в эволюции бактерий проиллюстрированы рассмотрением кишечные грамотрицательные бактерии и строение их плазмид. Бактерии собранные в доантибиотическую эру, содержали много плазмид, но они обычно отсутствовали детерминанты устойчивости. Многие из плазмид R из текущего клинические изоляты принадлежат к тем же группам несовместимости, что и обнаруженные плазмиды ранее, но они также определяют устойчивость к нескольким антибиотикам.В близкие отношения между их репликонами убедительно доказывают, что многие текущие плазмиды R произошли от более старых плазмид путем приобретения устойчивости детерминанты. Некоторые из плазмид с множественной устойчивостью к антибиотикам имеют индивидуальные транспозоны с несколькими детерминантами устойчивости, другие имеют несколько транспозоны резистентности расположены на разных участках, а третьи содержат сложные гибридные транспозоны устойчивости, образованные путем интеграции одного транспозона в другой. Поэтапное получение детерминант сопротивления может привести к в некоторых случаях к образованию составных транспозонов, кодирующих несколько детерминанты устойчивости.Терапевтическое использование антибиотиков и их включение в корм для животных обеспечивает селективные преимущества для бактерий с плазмидами R, тогда как конъюгация, трансформация и трансфекция обеспечивают средства для распространение плазмид R внутри и между видами бактерий. После плазмида, несущая транспозон, вводится в нового бактериального хозяина, транспозон и его детерминанты могут прыгать в хромосому или коренные плазмиды нового хозяина. Следовательно, стабильность мобилизующей плазмиды в новый бактериальный хозяин не важен для сохранения генетических детерминант расположен на транспозоне.
Рекомбинационная ДНК и клонирование генов
Существует множество методов создания гибридных молекул ДНК in vitro (рекомбинантная ДНК) и охарактеризовать их. Такие методы включают выделение определенных генов в гибридных репликоны, определяя их нуклеотидные последовательности и создавая мутации в обозначенные места (сайт-направленный мутагенез). Клон — это популяция организмы или молекулы, полученные в результате бесполого размножения от одного предка. Ген клонирование — это процесс включения чужеродных генов в репликоны гибридной ДНК.Клонированные гены могут быть экспрессированы в соответствующих клетках-хозяевах, и фенотипы, которые они определяют, что могут быть проанализированы. Некоторые ключевые концепции, лежащие в основе репрезентативных методов резюмируются здесь.
Первым шагом в клонировании генов является получение фрагментов донорской ДНК механическим или ферментативные методы. Некоторые эндонуклеазы рестрикции, обозначенные как класс II, являются особенно полезно для получения определенных фрагментов молекул ДНК. Они рассекаются обе нити двухцепочечных молекул ДНК в определенных палиндромных последовательностях (сайты рестрикции), длина которых обычно варьируется от четырех до восьми нуклеотидов, и полученные фрагменты ДНК называются рестрикционными фрагментами.Некоторое ограничение эндонуклеазы расщепляют совпадающие сайты с образованием фрагментов ДНК с тупыми концами, и другие разрезают в шахматном порядке для создания фрагментов ДНК с короткими, самокомплементарные одноцепочечные 5 ‘или 3’ концы (см.). Случайная вероятность того, что n соседние нуклеотиды в цепи ДНК будут соответствовать определенному сайту рестрикции составляет примерно 1/4 n . Сайты для ферментов, распознающих уникальные 4, 6 или 8 нуклеотидные мишени могут встречаться примерно один раз из каждых 256, 4096 или 65 536 нуклеотиды соответственно.Выбирая подходящие рестрикционные ферменты, специфическая ДНК молекулы, включая бактериальные хромосомы, плазмиды и геномы фагов, могут быть переваривается в наборы ограничивающих фрагментов, которые имеют соответствующие размеры для конкретных Приложения.
Таблица 5-3
Особенности ограничения типичного класса II Эндонуклеазы.
Карта ограничений определяет позиции целевых сайтов для конкретного ограничения эндонуклеазы в молекуле ДНК. Карты ограничений доступны для многих клонированных ДНК. фрагментов, плазмид и геномов фагов, а также для всей хромосомы E.coli и несколько других бактерий.
Второй этап клонирования генов — создание гибридных репликонов, состоящих из донорской ДНК. фрагменты и вектор клонирования (). Клонирующие векторы представляют собой небольшие репликоны плазмид или фагов, которые имеют один или несколько сайты рестрикции, в которые может быть вставлена чужеродная ДНК. Гибридные репликоны производится с использованием ДНК-лигазы для соединения рестриктированной векторной ДНК с донорной ДНК фрагменты, которые имеют совместимые концы, или, альтернативно, синтетические олигонуклеотиды используются в качестве линкеров для создания совместимости между донорскими и векторными молекулами ДНК с разными концами.Лигирование вектора с гетерогенным набором фрагментов ДНК из донорский геном называется клонированием дробовика, а сбор рекомбинантной ДНК молекулы, содержащие различные фрагменты, называются геномной библиотекой. Если специфический фрагмент ДНК доступен, он может быть включен в рекомбинантный репликон путем прямого клонирования в соответствующий вектор, выбранный из широкого разнообразия доступных векторов. Плазмидные и фаговые векторы используются в основном для клонирования небольших вставляет обычно меньше 10 кб. Примеры других векторов специального назначения включают космиды, которые представляют собой плазмидные векторы, которые могут быть упакованы в фаговые капсиды (лямбда космиды принимают вставки размером до 30-40 т.п.н.) и фагмиды, которые являются плазмид-фагами. гибридные репликоны, которые могут существовать в виде плазмид или одноцепочечных ДНК-фагов в разных условиях эксперимента.Космиды фага P1 могут принимать вставки до 100 т.п.н., а молекулы ДНК еще большего размера можно клонировать в дрожжевые искусственные хромосомы (YAC), которые могут стабильно поддерживать вставки размером до и более 1 Мбит / с в размер. Другие специализированные векторы обнаруживают промоторы, сигналы терминации транскрипции, или другие регуляторные элементы во вставках чужеродной ДНК или, наоборот, обеспечивают промоторы, с которых может быть инициирована транскрипция клонированных генов.
Рисунок 5-12
Схематическое изображение эксперимента по клонированию гена.Плазмидный вектор клонирования pBR322 имеет размер 4,36 пар оснований, имеет гены на устойчивость к ампициллину (amp r ) и тетрациклину ( tet r ) и имеет только один HindIII сайт рестрикции, расположенный внутри (подробнее …)
Заключительными этапами клонирования генов является введение гибридных репликонов в соответствующие клетки-реципиенты и проверить их на экспрессию интересующих донорских генов. Прокариотический клетки (включая бактерии) или эукариотические клетки (включая дрожжи, животные или растения) ячеек) могут использоваться в качестве получателей, но они различаются по своему вседозволенность для определенных репликонов, транскрипционные сигналы, которые они распознают и посттрансляционные модификации белковой структуры, которые они может выполнить.Можно вводить рекомбинантные молекулы ДНК, полученные in vitro. непосредственно в клетки-реципиенты путем трансформации или трансфекции. Кроме того, клоны в космидных или фаговых векторах могут быть упакованы в фаговые оболочки и введены в восприимчивые клетки-реципиенты путем трансдукции. Используя специализированные векторы (челнок векторов), которые могут реплицироваться в нескольких типах клеток, гены любого организма могут быть клонированы и обработаны в удобной бактериальной системе, а затем повторно введены в клетки исходного организма для анализа в их естественных окружающая обстановка.
Существует множество методов идентификации бактерий, содержащих рекомбинантную ДНК. молекулы. Большинство векторов клонирования имеют гены признаков, которые могут быть положительно выбран, например, устойчивость к антибиотикам. Кроме того, часто можно ввести чужеродную ДНК в вектор клонирования в сайт, который инактивирует несущественная, но легко узнаваемая векторная функция. Если оба эти условия выполняются, бактерии, содержащие рекомбинантные молекулы, могут быть отобраны и легко отличить от бактерий, содержащих только вектор.Бактерии в геномная библиотека, содержащая конкретный клонированный ген, может быть идентифицирована с помощью биохимические или иммунологические методы тестирования желаемого генного продукта. В качестве альтернативы интересующий клонированный ген может быть обнаружен непосредственно с помощью нуклеиновой кислоты. методы кислотной гибридизации при условии, что доступен конкретный ДНК- или РНК-зонд. Поскольку вставка чужеродной ДНК в вектор клонирования в соответствующем сайте делает не инактивировать его способность реплицироваться в соответствующих клетках-реципиентах, гибрид представляющие интерес репликоны могут быть амплифицированы путем репликации, и рекомбинантная ДНК молекулы или их генные продукты могут быть очищены и изучены.Способность очищать специфические молекулы ДНК сделали возможным развитие ферментативных и химических методов для определения их нуклеотидных последовательностей и современные методы введения мутации в определенных сайтах в клонированных генах основаны на знании их ограничения карты или нуклеотидные последовательности.
Методы рекомбинантной ДНК позволяют клонировать определенные фрагменты ДНК из любых источника в векторы, которые могут быть изучены на хорошо охарактеризованных бактериях, в эукариотические клетки, или in vitro .Применения клонирования ДНК быстро расширяется во всех областях биологии и медицины. В медицинской генетике такие область применения варьируется от пренатальной диагностики наследственных заболеваний человека до характеристика онкогенов и их роль в канцерогенезе. Фармацевтическая приложения включают крупномасштабное производство из клонированных человеческих генов биологических продукты с терапевтической ценностью, такие как полипептидные гормоны, интерлейкины и ферменты. Приложения в общественном здравоохранении и лабораторной медицине включают разработки вакцин для предотвращения конкретных инфекций и зондов для диагностики конкретных инфекции путем гибридизации нуклеиновых кислот или полимеразной цепной реакции (ПЦР).В в последнем процессе используются олигонуклеотидные праймеры и ДНК-полимераза для амплификации специфических последовательности ДНК-мишени во время нескольких циклов синтеза in vitro, что делает его возможно обнаружение редких целевых последовательностей ДНК в клинических образцах с большим чувствительность.
Регуляция экспрессии генов
Фенотипические свойства бактерий определяются их генотипами и ростом условия. Для бактерий в чистой культуре часто возникают изменения условий роста. в предсказуемых физиологических адаптациях у всех членов популяции.Как правило, основные генные продукты производятся в количествах, обеспечивающих самый быстрый рост данной среде, и продукты, необходимые при особых обстоятельствах, производятся только тогда, когда они нужны.
Физиологические адаптации часто связаны с изменениями метаболической активности. Поток метаболитов через определенные биохимические пути можно контролировать. как путем регулирования синтеза определенных ферментов, так и путем изменения активности существующих ферментов. Механизмы, регулирующие экспрессию генов, воздействуя на здесь обсуждается синтез конкретных генных продуктов.
Специфическая регуляция включает ген или группу генов, участвующих в конкретном метаболический процесс. Индукция и репрессия позволяют бактериям регулировать производство конкретных генных продуктов в ответ на соответствующие сигналы. Обычно катаболический ферменты индуцируются, когда субстрат для пути роста присутствует в ростовой среда, и биосинтетические ферменты подавляются продуктом пути. Ферменты, которые участвуют в одном биохимическом пути, часто занимают соседние положения на бактериальной хромосоме и координированно индуцируются или подавляются.Они образуют оперон, группу смежных генов, которые транскрибируются как единое целое. единица и переводится для получения соответствующих генных продуктов. Организация в оперон — важная стратегия для скоординированной регуляции экспрессии генов в бактериях. Опероны, которые могут быть индуцированы или подавлены, контролируются связыванием специфические регуляторные белки к определенным нуклеотидным последовательностям, которые функционируют как регуляторные сайты внутри оперона. Сравнение аминокислотных последовательностей многих этих различных регуляторных белков показали, что они могут быть сгруппированы вместе в семейства регуляторов (например,г. семейство белков lysR), которые могли развиться от общих предковых генов. Члены семейства lysR включают регуляторы таких разнообразные явления, такие как метаболизм лизина, цистеина и метионина у E. coli и репрессия железа в V. cholerae .
Глобальная регуляция одновременно изменяет экспрессию группы генов и оперонов, вместе называемые регулонами, которые контролируются одним и тем же регуляторным сигналом. Глобальное регулирование определяет реакцию бактерий на основные питательные вещества, такие как углерод, азот или фосфат, реакции на стрессы, такие как повреждение ДНК или тепло шока, а также синтез патогенами специфических факторов вирулентности при росте в их животные-хозяева.Количество определенного белка в бактериальной клетке может варьироваться. от нуля до многих тысяч молекул. Этот широкий диапазон часто определяется комбинированное действие нескольких регуляторных механизмов, влияющих на экспрессию соответствующий структурный ген. Регулирование достигается путем определения того, как часто ген транскрибируется в функциональную мРНК, насколько эффективно транслируется мРНК в белок, как быстро разлагается мРНК, как быстро белковый продукт переворачивается, и может ли активность белкового продукта быть изменена аллостерические эффекты или ковалентные модификации.
мРНК как транскрипционные единицы
Экспрессия гена начинается с ДНК-зависимой РНК-полимеразы (РНК-полимеразы) катализирует транскрипцию специфической мРНК из одной цепи ДНК-матрицы. Связывание РНК-полимеразы с ДНК происходит в определенных сайтах, называемых промоторами, и транскрипция начинается рядом с промотором. Сильные промоутеры могут взаимодействовать эффективно с РНК-полимеразой и инициировать транскрипцию с высокой скоростью; слабый промоторы инициируют транскрипцию с медленной скоростью. В любом случае мРНК синтезируется от его 5′-конца к его 3′-концу на приблизительно с постоянной скоростью, пока РНК-полимераза не распознает другой специфический сайт назвал терминатором.Затем РНК-полимераза отделяется от матрицы и транскрипция мРНК завершена.
Отдельные молекулы мРНК могут кодировать один или несколько полипептидов. Транскрипция оперон продуцирует полицистронную мРНК, которая кодирует несколько полипептидов. Трансляция полицистронных мРНК приводит к координированному синтезу кодируемых полипептиды, но каждый полипептид синтезируется как отдельная молекула. А специфический сайт связывания рибосомы расположен прямо перед началом каждого кодирующая последовательность на молекуле мРНК.
Информационные РНК в бактериях быстро разлагаются со средним периодом полураспада несколько минут, в отличие от тРНК и рРНК, которые намного более стабильны. Хотя мРНК составляют около половины вновь синтезированной РНК, они представляют собой только небольшая часть общей РНК. Короткий период полужизни мРНК имеет важные последствия для экспрессии генов. Если синтез определенной мРНК предотвращается, продукция соответствующих полипептидов быстро снижается.
Контроль экспрессии генов осуществляется путем регулирования одного или нескольких этапов путь от матрицы ДНК к продукту активного гена.Одновременный регулирование на нескольких уровнях позволяет лучше контролировать экспрессию генов, чем было бы возможно с помощью единого регулирующего механизма. Самый распространенный способ регулировать экспрессию генов в бактериях — контролировать производство специфических мРНК. Поскольку скорость удлинения молекулы РНК приблизительно равна константа, основными факторами, контролирующими синтез мРНК, являются скорость инициации и вероятность того, что полный текст будет произведено.
Регуляция инициации транскрипции
Некоторые мРНК в бактериях синтезируются с постоянной скоростью, что приводит к конститутивная продукция кодируемых полипептидов.Количество конкретных мРНК и полипептиды, продуцируемые разными конститутивными генами, сильно различаются, однако и часто отражают различия в силе сторонников тех гены.
Транскрипция многих оперонов регулируется в ответ на изменение окружающей среды. условия. Промоторы определяют максимальную скорость инициации транскрипции. для таких оперонов, но регуляторные белки участвуют в контроле транскрипция. Нуклеотидные последовательности в оперонах, к которым относятся специфические регуляторные связывание белков называется регуляторными сайтами или операторами.Операторы и промоутеры расположены близко друг к другу внутри оперонов и могут иметь перекрывающиеся ДНК последовательности. Связывание регуляторных белков с операторами может увеличиваться (положительное регулирование) или уменьшение (отрицательное регулирование) частоты инициация транскрипции. Белки, которые функционируют как негативные регуляторы: обычно называемые репрессорами. Поскольку регуляторные белки могут диффундировать через цитоплазма, структурные гены регуляторных белков не обязательно должны быть связаны к целевым оперонам.
Способность определять присутствие или отсутствие определенных соединений и изменять темпы синтеза соответствующих генных продуктов имеют центральное значение для контроля экспрессия гена. Регуляторные белки предлагают одно решение этой проблемы связь стимул-реакция. Многие регуляторные белки бифункциональны и связываются не только к соответствующим операторам, но и к конкретным эффекторам, которые небольшие молекулы, такие как определенные сахара, аминокислоты и другие метаболиты. Кроме того, регуляторные белки являются аллостерическими, что означает, что они могут существовать в различные конформации, которые проявляют различную аффинность связывания для своих родственные операторы и эффекторы.Достаточная концентрация эффекторных благовоний образование регуляторного комплекса белок-эффектор, который имеет либо высокий, либо низкое сродство к оператору в каждом конкретном случае. В отрицательно регулируемом систем, эффектор функционирует как корепрессор, если регулирующий белок-эффекторный комплекс является активным репрессором, а эффекторный комплекс функционирует как индуктором и вызывает дерепрессию, если свободный регуляторный белок является активным репрессор. Напротив, в положительно регулируемых системах эффектор стимулирует экспрессия оперона, если регуляторный белок-эффекторный комплекс представляет собой положительный регулятор, а эффектор подавляет экспрессию оперона, если свободный регуляторный белок является положительным регулятором.
Лактозный (lac) оперон E. coli является примером индуцибельный, отрицательно регулируемый оперон (). Ген lacI кодирует репрессор, который связывается с оператором lac и предотвращает транскрипцию из Промотор lac . Структурный ген этого репрессора отдельно от lac-оперона, и репрессор синтезируется конститутивно при низкая ставка. Когда индуктор связывается с lac-репрессором, комплекс не может связываться с оператора и не может предотвратить связывание РНК-полимеразы с промотором.При других благоприятных условиях экспрессируется оперон lac , в результате синтеза β-галактозидазы, β-галактозида пермеаза и β-галактозид трансацетилаза. Модель lac оперон может быть индуцирован лактозой или структурно родственными соединениями, такими как изопропил-b-D-тиогалактозид (IPTG). IPTG называют безвозмездным индуктором потому что он индуцирует lac-оперон, но не является субстратом для β-галактозидаза. Отрицательная регуляция также встречается во многих биосинтетических оперонов в E.coli . В таких оперонах продукт биосинтетический путь действует как эффектор для негативных регуляторных система.
Рисунок 5-13
Регулирование оперона lac в E. coli . Структурные гены lacZ , lacY и код lacA для β-галактозидазы, β-галактозид пермеаза и β-галактозид трансацетилаза, соответственно. Физиологическая роль lacA заключается в неизвестный. Репрессор lac (подробнее…)
Оперон арабинозы ( ara ) в E. coli является как положительно и отрицательно регулируется. В присутствии арабинозы нормативный белок стимулирует транскрипцию оперона ara. В отсутствие арабинозы однако регуляторный белок репрессирует оперон ara .
Опероны часто управляются более чем одним механизмом. Когда E. coli выращивают в среде, содержащей глюкозу и альтернативный источник углерода, такой как лактоза или арабиноза, индукция lac или ara оперон и использование лактоза или арабиноза задерживаются до тех пор, пока не будет израсходована глюкоза.Этот явление называется диуксическим ростом. Неспособность вызвать оперон lac или ara в присутствии глюкозы является примером репрессии катаболита. Лак и ara опероны положительно регулируются циклический-3 ‘, 5’-аденозинмонофосфат (цАМФ) и белок активатора гена катаболита (CAP) (продукт гена crp). В Комплекс цАМФ-ЦАП взаимодействует с сайтами связывания ЦАП в регуляторных областях некоторые опероны, в том числе опероны lac и ara, и стимулирует транскрипцию от соответствующих промоутеров.Уровень внутриклеточного цАМФ у E. coli высокий во время роста в отсутствие глюкозы и низкий во время роста в присутствии глюкозы. Катаболиты репрессии должны, следовательно, к отсутствию активации цАМФ-зависимых оперонов, когда бактерии выращены в присутствии глюкозы или другого быстро метаболизируемого углерода источники.
Регуляция терминации транскрипции
Аттенуация — это механизм регуляции оперонов путем прекращения транскрипции мРНК преждевременно.Аттенуация часто встречается в биосинтетических оперонах, включая trp , гистидин ( his ), треонин ( th ), изолейцин-валин ( ilv ) и фенилаланиновые (phe) опероны. Оперон trp в E. coli контролируется как репрессией, так и ослаблением. в наличие избытка триптофана, инициация транскрипции с trp Промотор репрессирован. Кроме того, однако, те транскрипты, инициированные с промотора trp , являются обычно заканчивается раньше любого из структурных генов Расшифровано trp, оперонов.Концентрация внутриклеточный триптофан, необходимый для поддержания репрессии, превышает уровень, необходимый для затухание. Такой двойной контроль позволяет клетке точно настроить выражение оперон trp в ответ на снижение концентрации триптофан.
Вторичная структура мРНК играет важную роль в механизме затухание. Все мРНК имеют лидерную последовательность между началом транскрипции. сайт и начало кодирующей последовательности для первого структурного гена.Для аминокислотные биосинтетические опероны, которые подвергаются аттенуации, лидер мРНК Последовательность имеет две отличительные особенности. Он кодирует короткий пептид, содержащий аминокислота, продуцируемая регулируемым путем, и она может образовывать альтернативу, взаимно несовместимые, двухцепочечные структуры РНК, которые участвуют в нормативные события. Например, пептид, кодируемый trp Лидерная последовательность мРНК содержит два соседних остатка триптофана, а пептид кодируемая лидерной последовательностью мРНК , имеет серию из семи последовательные остатки гистидина.показывает оперон trp и иллюстрирует альтернативный вторичный структур в лидерной последовательности trp мРНК . Есть три возможные вторичные структуры для этого региона, называемые сайтом паузы (сегменты 1 + 2), антитерминатор (сегменты 2 + 3) и аттенюатор (сегменты 3 + 4). Сегмент 1 сайта паузы перекрывается кодировкой область для пептида trpL . Какие второстепенные конструкции формируется зависит от эффективности трансляции trpL пептид.Когда сегменты 1 и 2 расшифровываются, они немедленно отжигаются и вызвать временную остановку РНК-полимеразы. Последующее инициирование трансляция пептида trpL нарушает сайт паузы и позволяет РНК-полимеразе продолжать транскрипцию. Если присутствует триптофан, транскрипция сегментов 3 и 4 и формирование структуры аттенюатора происходит, когда рибосома блокирует сегмент 2, в результате чего РНК-полимераза прекратить транскрипцию. Однако при дефиците триптофана триптофанил-тРНК также недостаточен, и рибосома останавливается на кодонах триптофана в сегменте 1.Это позволяет сегменту 2 отжигаться с вновь синтезированным сегментом 3, чтобы сформировать антитерминатор, тем самым делая сегмент 3 недоступным для отжига с сегментом 4. Таким образом предотвращается образование аттенюатора, и РНК-полимераза транскрибирует весь оперон trp . Таким образом истощение триптофан (фактически поставка триптофанил-тРНК) связан с регуляцией транскрипции биосинтетического оперона триптофана.
Рисунок 5-14
Регулировка оперона trp в E.coli . Организация оперона trp показана на верх фигуры. Пять структурных генов trpE , trpD , trpC , trpB и trpA кодируют ферменты которые катализируют терминальную последовательность реакций в триптофане формирование. (подробнее …)
В E. coli транскрипция и трансляция функционально спаренный. Бессмысленные мутации, вызывающие преждевременное прекращение трансляции часто вызывают снижение транскрипции более дистальных генов в том же опероне.Это явление называется полярностью. Рибосомы обычно инициируют трансляцию рост молекулы мРНК до завершения транскрипции, и такая трансляция маскирует сайты, которые в противном случае привели бы к прекращению действия РНК-полимеразы транскрипция. Преждевременное прекращение трансляции бессмысленным кодоном отделяет рибосомы от мРНК и позволяет РНК-полимеразе взаимодействовать с немаскированными сайтами терминации транскрипции.
В некоторых биологических системах, включая фаг лямбда, антитерминация используется как положительный регуляторный механизм для контроля экспрессии генов.Незамедлительно после заражение E. coli лямбда, РНК-полимераза связывается с двумя промоторов в лямбда-ДНК и инициирует дивергентные первичные транскрипты, которые оканчиваются на определенных сайтах лямбда-генома. Белок, кодируемый одним из первичные транскрипты взаимодействуют с РНК-полимеразой и позволяют ей продолжать транскрипция через первичные сайты терминации, тем самым выражая второй набор лямбда-генов. Один из продуктов, закодированный вторичной расшифровкой блокирует терминацию другой мРНК и активирует экспрессию третьего набора гены.Следовательно, антитерминация играет ключевую роль в управлении каскадом экспрессия гена во время литического роста фага лямбда. Антитерминация также участвует в регуляции оперонов рРНК E. coli.
Регуляция трансляции
Сайт связывания рибосомы на мРНК комплементарен последовательности на 3′-конец 16S рРНК. Взаимодействие между этими последовательностями облегчает формирование комплекса инициации синтеза белка. Как степень гомология с 16S рРНК и расстояние сайта связывания рибосомы от кодон инициации влияет на эффективность инициации трансляции.Использование кодонов в мРНК также влияет на эффективность трансляции. Информационные РНК для белков, которые требуются в больших количествах, как правило, используют кодоны, которые транслируются наиболее многочисленные виды тРНК, и обратное тоже верно.
Трансляционный контроль важен для регуляции синтеза рибосомных белки. Производство рибосом связано с высокими метаболическими затратами для бактерий, и при высоких скоростях роста рибосомы могут составлять почти половину клетки. масса. Большинство рибосомных белков и рРНК собраны в рибосомы, и пул свободных субъединиц рибосом очень мал.Гены рибосом белки организованы в несколько оперонов. Некоторые из свободных рибосом белки напрямую ингибируют трансляцию полицистронных мРНК, которые кодируют их, тем самым обеспечивая баланс синтеза рибосомальных белков с требование к их утилизации.
Регулоны и белки, передающие сигнал
Реглон — это группа генов или оперонов, контролируемых общим регулятором. Там есть несколько преимуществ в том, чтобы поставить под контроль разные опероны одним и тем же регулятор.Это позволяет связать восприятие одиночного стимула с экспрессия большого количества генов, которые могут быть необходимы для соответствующего ответ, и это устраняет потребность в скоординированно регулируемых генах быть сцепленным на бактериальной хромосоме. Стимул, к которому регулон Ответы могут быть внутриклеточным компонентом или сигналом окружающей среды. Отдельные опероны также могут регулироваться несколькими различными механизмы и выражаются в условиях, отличных от тех, которые влияют на весь регулон.
Более 40 различных регулонов были идентифицированы в E. coli . Конкретные примеры регулонов, которые отвечают на внутриклеточные компоненты, включают: описанный ранее регулон цАМФ-CAP и регулоны, контролируемые строгий ответ и ответ SOS. Когда рибосомы сталкиваются с незаряженной тРНК молекул во время синтеза белка активируется строгий ответ и приводит к быстрому прекращению синтеза рРНК. Новый нуклеотид под названием гуанозин-3′-дифосфат-5′-дифосфат (ppGpp) накапливается во время аминокислотного голодания.PpGpp, продуцируемый холостыми рибосомами, кажется, быть посредником строгого ответа, но точный механизм, вызывающий ингибирование синтеза рРНК неизвестно. Ответ SOS связан с повреждение ДНК и включает индукцию более 20 генов, участвующих в нескольких Пути репарации ДНК. Продукт гена recA обнаруживает ингибирование ДНК синтез и инициирует события, ведущие к протеолитическому расщеплению и инактивации репрессора пути SOS, кодируемого lexA ген.
Некоторые регулоны индуцируются определенными раздражителями окружающей среды, например, питательными веществами. ограничение или осмотический стресс. Часто опероны более чем одного регулона могут быть индуцированный, а термин «стимулон» использовался для описания набора генов, так что индуцированный. Обычно бактерии воспринимают такие условия окружающей среды двумя способами. компонентные системы. Первый компонент — это трансмембранный белок с внеклеточные и внутриклеточные домены. Его внеклеточный домен обнаруживает стимул окружающей среды, и его цитоплазматический домен передает сигнал.В второй компонент — бифункциональный цитоплазматический белок. Имеет домен-получатель который взаимодействует с модулем передатчика первого компонента, а также с эффекторный домен, контролирующий экспрессию соответствующего регулона. В модули передатчика и приемника двухкомпонентных систем регулирования от большое количество регулонов генетически связаны и имеют гомологию аминокислот. Сигнально-детектирующий и эффекторный домены белков из разных Однако регулоны различаются и определяют детектируемый сигнал и опероны, которые активируются или подавляются в ответ на этот сигнал.
Глобальное регулирование играет важную роль в физиологии патогенных бактерий. Например, Vibrio cholerae и Bordetella. pertussis выражают многие из детерминант вирулентности в управление системами передачи сигналов, относящимися к двухкомпонентной системы, описанные выше. Экспрессия белков, необходимых для инвазивного Фенотип контролируется температурой в Shigella . Yersinia enterocolitica чувствует температуры и концентрации ионов кальция и связывает эти сигналы с экспрессия генов и клеточная локализация продуктов генов, которые подходит для внутриклеточной или внеклеточной среды.В тканях хозяина концентрация свободного железа крайне низка, и большинство болезнетворных бактерий имеют системы транспорта железа с высоким сродством, которые индуцируются при низком уровне железа условия. Синтез дифтерийного токсина C. diphtheriae , токсин Shiga by Shigella dysenteriae , экзотоксин А от Pseudomonas aeruginosa и другие специфические белков у многих патогенных бактерий индуцируется в условиях ограниченного железом рост. Эти примеры показывают, как факторы окружающей среды могут регулировать экспрессия генов вирулентности у патогенных бактерий.
Бактерии | Что такое микробиология?
Бактерии — одноклеточные микробы. Структура клетки проще, чем у других организмов, поскольку в ней нет ядер или мембраносвязанных органелл. Вместо этого их центр управления, содержащий генетическую информацию, находится в единой петле ДНК. У некоторых бактерий есть дополнительный круг генетического материала, называемый плазмидой. Плазмида часто содержит гены, которые дают бактерии некоторое преимущество перед другими бактериями. Например, он может содержать ген, который делает бактерии устойчивыми к определенному антибиотику.
Бактерии подразделяются на пять групп в зависимости от их основных форм: сферические (кокки), стержневые (бациллы), спиральные (спириллы), запятые (вибрионы) или штопоры (спирохеты). Они могут существовать в виде отдельных ячеек, в парах, цепочках или кластерах.
© ttsz / iStock Различные формы бактерий.Бактерии встречаются во всех средах обитания на Земле: в почве, камнях, океанах и даже в арктическом снегу. Некоторые живут внутри или на других организмах, включая растения и животных, включая людей.В организме человека примерно в 10 раз больше бактериальных клеток, чем клеток человека. Многие из этих бактериальных клеток выстилают пищеварительную систему. Некоторые бактерии живут в почве или на мертвых растениях, где они играют важную роль в круговороте питательных веществ. Некоторые виды вызывают порчу продуктов питания и повреждение урожая, но другие невероятно полезны при производстве ферментированных продуктов, таких как йогурт и соевый соус. Относительно немного бактерий являются паразитами или патогенами, вызывающими болезни у животных и растений.
© Гаэтан Стоффель / iStock Трехмерная иллюстрация Escherichia coliКак размножаются бактерии?
Бактерии размножаются двойным делением. В этом процессе бактерия, представляющая собой единственную клетку, делится на две идентичные дочерние клетки. Бинарное деление начинается, когда ДНК бактерии делится на две части (повторяется). Затем бактериальная клетка удлиняется и разделяется на две дочерние клетки, каждая из которых имеет ДНК, идентичную родительской клетке.Каждая дочерняя клетка является клоном родительской клетки.
При благоприятных условиях, например при правильной температуре и наличии питательных веществ, некоторые бактерии, такие как Escherichia coli , могут делиться каждые 20 минут. Это означает, что всего за семь часов одна бактерия может произвести 2 097 152 бактерии. Еще через час количество бактерий вырастет до колоссальных 16 777 216 человек. Вот почему мы можем быстро заболеть, когда в наш организм вторгаются патогенные микробы.
Механизм выживания
Некоторые бактерии могут образовывать эндоспоры.Это спящие конструкции, которые чрезвычайно устойчивы к агрессивным физическим и химическим условиям, таким как тепло, УФ-излучение и дезинфицирующие средства. Это очень затрудняет их уничтожение. Многие бактерии, продуцирующие эндоспоры, являются опасными патогенами, например Bacillus anthracis , вызывающая сибирскую язву.
Микробиология сегодня: микобактерии
Микробактерии, вызывающие туберкулез у людей и животных, Mycobacterium tuberculosis и Mycobacterium bovis , описаны в этом выпуске журнала Microbiology Today наряду с Mycobacterium leprae , вызывающей проказу, и cerabacterium. Язва Бурули.
Строим бактериальные мостики
Часто первое, что приходит на ум, когда мы думаем о микробах в искусственной среде, — это повреждение, разложение, изменение цвета и окрашивание строительных материалов и их поверхностей. То, что мы часто не принимаем во внимание, — это их способность действовать как «биоинженеры».
Объяснение туберкулеза
Туберкулез (ТБ) — изнурительное полиорганное заболевание, вызываемое бактерией Mycobacterium tuberculosis.Самая серьезная форма заболевания — туберкулез легких, инфекция легких и дыхательных путей.
Устойчивость к противомикробным препаратам (УПП)
Угроза устойчивости к противомикробным препаратам (УПП) теперь признана во всем мире, и, по оценкам, 10 миллионов человек в год будут умирать из-за устойчивости к противомикробным препаратам к 2050 году, если не будут приняты срочные меры.
Псевдомонады — друг и враг
Виды рода Pseudomonas являются одними из наиболее изученных бактерий в научном сообществе.Бактерии этого рода широко используются в качестве модельных организмов в микробных исследованиях и включают ряд важных видов в таких областях, как патогенность растений, биоремедиация и микробиология окружающей среды.